<宇宙の歴史と地質時代>
Update 2024.09.05

TOP|
宇宙創成は138億年前:宇宙マイクロ波背景放射の温度は完全に一様ではなく、ごくわずかのムラがあることが発見され、
このムラを分析すると宇宙の年齢が判明した
宇宙の温度が下がって可視光より波長が長い電波になったのが宇宙マイクロ波背景放射
宇宙創成38万後の宇宙の晴れ上がり直後の姿が宇宙マイクロ波背景放射なのです
宇宙の晴れ上がり前の宇宙は光子は自由電子の雲の中で散乱して、濃い霧の状態で光によっては
観測できません(重力波による観測が期待されている)
約60億年前宇宙の加速膨張:ハッブルにより遠い銀河ほど赤方偏移が大きくなること(宇宙の膨張)が確かめられた後、
Ia型超新星を標準光源とする銀河までの距離と赤方偏移の測定によって、宇宙の膨張の歴史が
再度調べられた結果、宇宙の膨張速度はなぜか加速に転じていた
ダークエネルギーの斥力が物質の引力を上回り、宇宙の膨張速度が加速し始めた
太陽系誕生は46億年前:コンドライトという隕石の生成年代を調べてわかった
コンドライト隕石中の鉱物を鉛の放射性同位体206Pbと207Pbを使った年代測定を行い、隕石の中で
かなり早い時期に形成される鉱物の形成年代が45億6820万年前であると結論づけた
分子雲→原始星→主系列星の誕生(中心部で核融合反応の開始)
生命誕生は約40億年前:生物の進化をさかのぼり、全生物の最後の共通祖先(コモノート)は38億年前と推定された
グリーンランドのイスア地域の岩石に38億年前に活動していた生物の痕跡がグラファイトとして
残されていると報告、39億5000万年前より古い堆積岩のグラファイトにも痕跡がある事を発見した
6550万年前の大量絶滅:小惑星の衝突により、恐竜が絶滅した
イリジウムが凝集した粘土層→衝突クレーターの発見→天体衝突を裏付け→大量絶滅
人類の出現は700万年前:トゥーマイ猿人の出現
霊長類の進化→人類の出現→人類の進化:猿人→原人→旧人→新人
宇宙史年代
|TOP宇宙創成は138億年前:宇宙マイクロ波背景放射の温度は完全に一様ではなく、ごくわずかのムラがあることが発見され、
このムラを分析すると宇宙の年齢が判明した
宇宙の温度が下がって可視光より波長が長い電波になったのが宇宙マイクロ波背景放射
宇宙創成38万後の宇宙の晴れ上がり直後の姿が宇宙マイクロ波背景放射なのです
宇宙の晴れ上がり前の宇宙は光子は自由電子の雲の中で散乱して、濃い霧の状態で光によっては
観測できません(重力波による観測が期待されている)
約60億年前宇宙の加速膨張:ハッブルにより遠い銀河ほど赤方偏移が大きくなること(宇宙の膨張)が確かめられた後、
Ia型超新星を標準光源とする銀河までの距離と赤方偏移の測定によって、宇宙の膨張の歴史が
再度調べられた結果、宇宙の膨張速度はなぜか加速に転じていた
ダークエネルギーの斥力が物質の引力を上回り、宇宙の膨張速度が加速し始めた
太陽系誕生は46億年前:コンドライトという隕石の生成年代を調べてわかった
コンドライト隕石中の鉱物を鉛の放射性同位体206Pbと207Pbを使った年代測定を行い、隕石の中で
かなり早い時期に形成される鉱物の形成年代が45億6820万年前であると結論づけた
分子雲→原始星→主系列星の誕生(中心部で核融合反応の開始)
生命誕生は約40億年前:生物の進化をさかのぼり、全生物の最後の共通祖先(コモノート)は38億年前と推定された
グリーンランドのイスア地域の岩石に38億年前に活動していた生物の痕跡がグラファイトとして
残されていると報告、39億5000万年前より古い堆積岩のグラファイトにも痕跡がある事を発見した
6550万年前の大量絶滅:小惑星の衝突により、恐竜が絶滅した
イリジウムが凝集した粘土層→衝突クレーターの発見→天体衝突を裏付け→大量絶滅
人類の出現は700万年前:トゥーマイ猿人の出現
霊長類の進化→人類の出現→人類の進化:猿人→原人→旧人→新人
| 時刻 | 温度(K) | エネルギー(eV) | 宇宙の創成 物質相 力の分岐 |
|---|---|---|---|
| ~10-44秒:プランク時間 | - | - | 真空の量子ゆらぎ → 真空の相転移 |
| 10-44秒後:宇宙創成の瞬間 | 1032度:1京度の1京倍 | 1028eV:100兆eVの100兆倍 | 超々微小宇宙 重力の分離 |
| 10-36秒後:宇宙創成直後 | 1028度:1000兆度の10兆倍 | 1024eV:1兆eVの1兆倍 | 強い力の分離 |
| 10-36秒~10-34秒後 | 真空の相転移で生じる光速を超える指数関数的な膨張 | インフレーション(10-33cm→1033倍) | |
| 10-34秒後:ビッグバン | 真空のエネルギーで宇宙が再加熱された火の玉宇宙 | 物質(粒子・反粒子)と光の誕生 | |
| 10-11秒後:ビッグバン直後 | ~数千兆度 | ~1011eV:千億eV以上 | 超微小宇宙 弱い力・電磁気力の分離 |
| 10-12秒~10-6秒後 | ~5兆度 | ~5億eV | クォーク・グルオン相 ヒッグス機構 |
| 10-6秒~1秒後~3分後 | ~500億度 | ~500万eV | 核子・電子・ニュートリノ相:素粒子 |
| 3分~20分後 | ~50億度 | ~50万eV | 原子核相:重陽子・ヘリウム核 |
| 20分~10万年後 | ~4000度 | ~0.5eV | 原子相:原子核・電子 |
| 10万年~38万年後 | 自由電子が原子核に束縛され光子が直進(3000度程度) | 宇宙の晴れ上がり 電離→結合 | |
| 38万年~数億年後 | 漂う原子や自由な光子・ニュートリノだけの真暗宇宙 | 宇宙の暗黒時代 星の重力凝集前 | |
| 2億年~数億年後 | 最初の恒星群の誕生 → 鉄までの重い元素の生成 | 最初の星の誕生 宇宙の再電離 | |
| 10億年~30億年後 | → 超新星爆発 → 鉄よりも重い元素の生成 | 銀河の形成~銀河団の形成 | |
| 78億年後:60億年前 | ダークエネルギーによる宇宙の加速膨張 | 第2のインフレーション | |
| 92億年後:46億年前 | 軽い元素 → 星間ガス雲(分子雲) → 双極分子流 → 収縮進み核融合開始 → 原始太陽 | 原始太陽系円盤 → 太陽系の誕生 星間ガスが散逸 → 岩石惑星 星間ガスを取込 → 巨大ガス惑星 重力の大きさで相違 巨大氷惑星 | |
| 重い元素 → ダスト → 微惑星 → 衝突・合体 → 暴走成長 → 原始惑星 → 巨大衝突 | |||
| 46億年前~138億年後 | 冥王代→太古代→原生代→古生代→中生代→新生代 | 地質時代 → 先史時代 | |
| 138億年後 | 現在の宇宙 | 有史時代(歴史時代) | |
K:ケルビン(水の三重点温度0.01℃を273.16Kとする絶対温度),eV:電子ボルト(1eV=1電子を1ボルトで加速時のエネルギー)
104=1万,108=1億,1012=1兆,1016=1京,10-12=1兆分の1,1028=1015+13=1015×1013=1000兆×10兆=100兆×100兆
インフレーションの膨張速度は光速を超えていた:特殊相対性理論が禁じているのは「運動」であって空間の膨張ではない
ヒッグス機構 :宇宙が膨張して冷えてきたある時点で、ヒッグス粒子が凝縮して真空に満ち素粒子が動きにくくなった
宇宙背景ニュートリノ:宇宙創成数秒後、宇宙の膨張によりエネルギーや密度が下がりニュートリノは衝突せずに飛び去る
宇宙ニュートリノ背景放射:宇宙創成3分~20分後、原子核の合成(ビッグバン元素合成)の過程でニュートリノを放出
宇宙の再電離 :星からの強い紫外線の衝突によって宇宙に漂う水素原子の原子核と電子がばらばらに別れて分離された
|
──────────宇宙の相転移(力の分化)────────── プランク時間 宇宙(時空)が生まれる、時空の量子的ゆらぎの終わり 10-44秒後 第1の相転移:重力の分離、クォークの誕生 10-36秒後 第2の相転移:強い力(色)の分離、グルーオンの誕生 10-11秒後 第3の相転移:弱い力の分離(電弱分離)、電子の誕生 ゲージ対称性 ヒッグス場 光子だけ無質量・光速 電弱対称性─→対称性の自発的破れ─→電弱分離 質量を禁止 ヒッグス粒子の出現 弱ボソンが質量を獲得 ──────輻射の時代の終わり、物質の時代の始まり────── 10-4秒後 第4の相転移:クォーク(素粒子)がハドロン(核子)に 100秒後 軽元素合成の開始(核融合反応)、He,D,Liなどの合成 |
|
素粒子 ⇒ 核子 ⇒ 原子核 ⇒ 原子
クォーク(uud) … 陽子──┐
陽子は、 クォークからできている├→原子核─┐ クォーク(udd) … 中性子─┘ ├→原子 レプトン(e-) … 電子──┘ 原子核は、陽子と中性子からできている 原子は、 原子核と電子からできている |
|
色の力で、クォーク → 陽子 :3種の色で結びつく、色の力を伝えるのはグルーオン 弱い力で、陽子 → 中性子:β変換、陽子(uud)→中性子(udd) 核力で、 陽子+中性子 → 原子核:核内で陽子と中性子がπ中間子の放出・吸収、核力が発生 電気力で、原子核+電子 → 原子 :原子内で原子核と電子が光子の放出・吸収、電気力が発生 |
◆ビッグバンのあと、宇宙の温度がどんどん下がっていき、粒子が反粒子より多くなった経緯
当初は、粒子とともにペアの反粒子も対生成したが、すぐに対消滅したりをくりかえしていた … 対称:粒子数=反粒子数
10の-10乗秒後のどこかで、「対称性の自発的破れ」により、反粒子は4秒後までに消失する … 破れ:粒子数>反粒子数
そして、「対称性の自発的破れ」によって、わずかに多くなった粒子だけが残るようになった … 対生成が起こらなくなる
粒子:クォーク・ハドロン・レプトン、反粒子:反クォーク・反ハドロン・反レプトン、中間子は自分自身が反粒子である
※:「消えた反物質」の謎については、さまざまな機構が提案されているが、どれが正しいのか現在はまだ確定していない
→ 消えた反物質の謎の解明に挑む
|
|
ヘリウム4核:ヘリウムの同位体であるヘリウム4の原子核(陽子2個と中性子2個) … ビッグバンの3分後に大量生成
恒星内部での元素合成=水素燃焼反応(PPⅠ反応=Proton-ProtonⅠChain Reaction):現在の太陽活動の主要なエネルギー源
水素核融合反応は結局 4p → He4 + 2e+ + 2νe + 2γとなる反応である(e+:陽電子,νe:電子ニュートリノ,γ:光子)
陽子が中性子に変われるのは重水素の原子核の質量が、もともとの陽子2個の合計の質量よりも軽くなるからです
この現象は「質量欠損」と言い、軽くなるということはエネルギーが下がることなので、陽子が中性子に変われるのです
4つの陽子が核融合によりヘリウム4核になる際に、質量は0.7%ぐらい減り、その分の静止質量エネルギーに相当する2500
万eVの核エネルギーが解放され、エネルギーの一部は太陽の外に飛び出すニュートリノが持ち去り、そのほかのガンマ線の
エネルギーは、太陽内の電子と衝突を通して熱エネルギーとなり、10万年かけて太陽表面に達し、太陽光として放射される
+電荷の陽子同士の強い反発力に打ち勝つ陽子の激しい運動や反応温度未満でも起こるトンネル効果で核融合が起きている
恒星内部での水素燃焼には、PPⅠ反応とCNOサイクルの両方が働いているが、CNOサイクルは大質量星のエネルギー
生成過程に大きく寄与している→太陽内部でCNOサイクルによって生み出されるエネルギーは全体の約1.6%に過ぎない
※CNOサイクル:炭素(C)・窒素(N)・酸素(O)が円環状に連続した触媒核になって水素核(陽子)が燃える原子核反応
┌─────────────原始太陽の形成───────────┐
星間分子雲──→分子雲の収縮─→ガス円盤と双極分子流─→原始太陽
分子雲コアの収縮でできた原始太陽に落ち込むガスやダストが、回転しながら、周囲に平らな円盤を形成した
原始太陽の中心部が約1000万度に達すると、水素中心核融合反応が起こり、主系列星となって太陽が誕生した
┌─────────────────────原始惑星の形成─────────────────────┐
円盤の温度が下がるとガスは個体微粒子に凝縮、太陽に近い部分は岩石と金属に遠い部分は氷等の物質となる
ガス・個体微粒子→ダスト層→微惑星→衝突合体→原始惑星:内周部は岩石惑星、外周部は氷惑星→ガス捕獲
弱い重力の惑星は星間ガスが太陽風で吹き飛び、強い重力の惑星は星間ガスを捕獲して巨大氷惑星となった
星間分子雲──→分子雲の収縮─→ガス円盤と双極分子流─→原始太陽
| 星間物質 | 分子雲 | 分子雲コア | |
| 密度 | 1cm3水素原子数個 | まだら | 1cm3水素分子10万~100万個 |
| 大きさ | 100光年 | 0.1光年 | |
| 質量 | 太陽の10万倍 | 太陽の10倍 |
原始太陽の中心部が約1000万度に達すると、水素中心核融合反応が起こり、主系列星となって太陽が誕生した
┌─────────────────────原始惑星の形成─────────────────────┐
円盤の温度が下がるとガスは個体微粒子に凝縮、太陽に近い部分は岩石と金属に遠い部分は氷等の物質となる
ガス・個体微粒子→ダスト層→微惑星→衝突合体→原始惑星:内周部は岩石惑星、外周部は氷惑星→ガス捕獲
弱い重力の惑星は星間ガスが太陽風で吹き飛び、強い重力の惑星は星間ガスを捕獲して巨大氷惑星となった
岩石惑星 スノーライン 巨大ガス惑星 巨大氷惑星 短周期彗星源 長周期彗星源
地球型惑星 アステロイドベルト 木星型惑星 天王星型惑星 カイパーベルト 球殻状の微惑星群
太陽●→○─→○──→○──→○──→◎──→○──→○──→○──→○───→◎───→◎──∥──→◎
水星 金星 地球 火星 小惑星帯 木星 土星 天王星 海王星 冥王星 散乱円盤天体 オールトの雲
↑ ↑ ↑ ↑ ↑ ↑
└────太陽系内周部────┘ └──太陽系外周部───┘ └──太陽系外縁天体──┘
0 1 2 3 4 太陽からの
0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 0 1 2 3 4 5億km ← 平均距離
─+-+─────-+─────-+──────────────-+────────────-+-
地火 木 土 天 海
球星 星 星 王 王
星 星
地球型惑星 アステロイドベルト 木星型惑星 天王星型惑星 カイパーベルト 球殻状の微惑星群
太陽●→○─→○──→○──→○──→◎──→○──→○──→○──→○───→◎───→◎──∥──→◎
水星 金星 地球 火星 小惑星帯 木星 土星 天王星 海王星 冥王星 散乱円盤天体 オールトの雲
↑ ↑ ↑ ↑ ↑ ↑
└────太陽系内周部────┘ └──太陽系外周部───┘ └──太陽系外縁天体──┘
0 1 2 3 4 太陽からの
0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 0 1 2 3 4 5億km ← 平均距離
─+-+─────-+─────-+──────────────-+────────────-+-
地火 木 土 天 海
球星 星 星 王 王
星 星
┌──────太陽圏:太陽風の勢力圏(ヘリオスフィア)─────┐ 恒星間領域
│ ヘリオシース ヘリオポーズ
├────────────────┬…………………………………: 太陽風の圧力と星間物質や
│ →超音速の太陽風→ │ →亜音速の太陽風→ ≫⇔≪ ←銀河系の磁場の圧力が釣り合う
太陽●─∥→○────→◎─────→○────────────→●────∥───→◎
│ 海王星 カイパーベルト 終端衝撃波面 太陽圏の境界 オールトの雲
│ 太陽から30AU ~50AU↑ 75~90AU 太陽から123AU 太陽から1万~10万AU
└───────────────太陽系:太陽の重力圏───────────────┘
│ ヘリオシース ヘリオポーズ
├────────────────┬…………………………………: 太陽風の圧力と星間物質や
│ →超音速の太陽風→ │ →亜音速の太陽風→ ≫⇔≪ ←銀河系の磁場の圧力が釣り合う
太陽●─∥→○────→◎─────→○────────────→●────∥───→◎
│ 海王星 カイパーベルト 終端衝撃波面 太陽圏の境界 オールトの雲
│ 太陽から30AU ~50AU↑ 75~90AU 太陽から123AU 太陽から1万~10万AU
└───────────────太陽系:太陽の重力圏───────────────┘
AU:1天文単位=1億4959万7870.7km(地球と太陽の平均距離に由来)、123AU(太陽圏の境界)=184億54万km
太陽から噴き出す太陽風の勢力圏(太陽風の荷電粒子が存在する領域)を太陽圏(ヘリオスフィア)という
太陽風は太陽系外縁部で天の川銀河内の星間物質や星間磁場と衝突し、終端衝撃波(termination shock)を形成する
太陽から太陽風が100万km/h以上の速度で吹く、半径約150億kmの終端衝撃波面は、ほぼ球形です
太陽圏では、太陽風の外向きの圧力が、入ってくる星間物質の圧力と釣り合っています
太陽風が星間物質や銀河系の磁場と衝突して完全に混ざり合う境界面をヘリオポーズ(Heliopause)と言う
終端衝撃波面の外側は、低速度の太陽風と星間物質とが混ざり合うヘリオシースという領域です
カイパーベルトの分布範囲は、概ねヘリオポーズの内側にあり、オールトの雲は完全にヘリオポーズの外側にある
太陽圏の圧力で生じる星間物質乱流領域(バウショック)は星間物質の中を進む太陽の速度が遅いため形成されない
地球型惑星の星間ガスや原始大気上層の軽い水素は太陽風に吹き飛ばされた
原始惑星創成時のガス成分は、ほとんどが強い太陽風で吹き飛ばされた
火星にはかつて大気があったが、太陽風がそれを吹き飛ばしてしまった
地球型惑星の初期大気は、その後の火山活動で固体部分から放出された
大部分は、形成初期の激しい火山活動によって脱ガスとして放出された
この時点では、惑星内部に核が形成され、磁場が太陽風をさえぎった
磁気バリアに守られた地球の宇宙空間領域は「地球磁気圏」と呼ばれています
太陽風は、地上から3万km(静止軌道と呼ばれる高度付近)から10万km
くらいの位置で食い止められていて、その反対側の磁気圏は太陽風に吹
き流されたような形をしていることがわかっています
原始惑星創成時のガス成分は、ほとんどが強い太陽風で吹き飛ばされた
火星にはかつて大気があったが、太陽風がそれを吹き飛ばしてしまった
地球型惑星の初期大気は、その後の火山活動で固体部分から放出された
大部分は、形成初期の激しい火山活動によって脱ガスとして放出された
この時点では、惑星内部に核が形成され、磁場が太陽風をさえぎった
磁気バリアに守られた地球の宇宙空間領域は「地球磁気圏」と呼ばれています
太陽風は、地上から3万km(静止軌道と呼ばれる高度付近)から10万km
くらいの位置で食い止められていて、その反対側の磁気圏は太陽風に吹
き流されたような形をしていることがわかっています
| 地球の内側・外側 | 内惑星 | 外惑星 | ||||||
|---|---|---|---|---|---|---|---|---|
| 惑星 | 水星 | 金星 | 地球 | 火星 | 木星 | 土星 | 天王星 | 海王星 |
| 太陽からの平均距離 | 0.5790億km | 1.0820億km | 1億4960万km | 2.2790億km | 7.7830億km | 14.3億km | 28.7億km | 45億km |
| 惑星の大きさ | 0.4880万km | 1.2104万km | 1.2742万km | 0.6795万km | 14.2984万km | 12.537万km | 5.1119万km | 4.9529万km |
| 大きさの地球比 | 0.383 | 0.95 | 1 | 0.533 | 11.221 | 9.839 | 4.0119 | 3.887 |
| 公転周期 | 0.24 | 0.62 | 1 | 1.88 | 11.9 | 29.5 | 84.0 | 164.8 |
| 自転周期 | 58.646日 | 243日逆行 | 24時間 | 24.5時間 | 10時間 | 10時間 | 17時間逆行 | 16時間 |
| 質量の地球比 | 0.055倍 | 0.82倍 | 1 | 0.107倍 | 318倍 | 95.2倍 | 14.5倍 | 17.2倍 |
| 惑星の分類 | 地球型惑星 | 木星型惑星 | 天王星型惑星 | |||||
| 核 | 鉄・ニッケル合金(火星は+硫化鉄) | 岩石・氷 | ||||||
| マントル | ケイ酸塩 | 液体金属水素(ヘリウム含む) | アンモニア・水・メタン混合の氷 | |||||
| 地殻 | ケイ酸塩 | - | ||||||
| 大気 | - | 主に二酸化炭素 | 主に窒素と酸素 | 主に二酸化炭素 | 液体分子水素(気体を含む) | ヘリウム・メタンを含む水素ガス | ||
| 惑星の衛星数 | 0 | 0 | 1 | 2 | 72 | 66 | 27 | 14 |
海王星の最大の衛星であるトリトンは、太陽系で唯一、逆行軌道をたどり、惑星の自転と反対方向に移動する大きな衛星です
太陽からの距離:遠からず近からず程よく太陽エネルギーを受け取る距離、液体の水の存在、大気の組成、など
地球は、太陽から22億分の1のエネルギー(光や熱)を受け取って、その恩恵によって豊かな自然が育まれ多種
多様な生命が暮らす生命の星ですが、太陽から来るのはそれだけでなく太陽風と呼ばれる高温のプラズマがある
惑星の大きさ:惑星内部の活動が存在できる質量、太陽風の影響下でも大気を保つ磁場と重力がある
| |
水が蒸発してしまう領域 ← | ハビタブルゾーン | → 水が凍ってしまう領域
| |
太陽●────────→○────────→●────────→○────────→
金星 | 地球 | 火星
| |
| |
水が蒸発してしまう領域 ← | ハビタブルゾーン | → 水が凍ってしまう領域
| |
太陽●────────→○────────→●────────→○────────→
金星 | 地球 | 火星
| |
| |
「太陽風」は太陽系外から流入する銀河宇宙線をブロックして、銀河宇宙線の地球に対する影響を抑えています
太陽風自体も放射線をもっていますが、今度は「地磁気」がこれをおよそ完全にブロックしています
最後に「大気」が太陽風のすき間をぬって侵入してきた銀河宇宙線をブロックしています
| 月 | 地球 | 太陽 | 太陽系 | 天の川銀河 | ラニアケア超銀河団 | 観測可能な宇宙 |
|---|---|---|---|---|---|---|
| 3,474.8km | 12,742km | 1,392,700km | 3.15光年 | 10万5700光年 | 5億光年 | 930億光年 |
| 1万3千km | 139万km | 30兆km | 100京km | 50垓km | 9300垓km | |
| 3.5×103km | 1.3×104km | 1.4×106km | 3×1013km | 1×1018km | 5×1021km | 9.3×1023km |
| 地球の4分の1 | 1 | 地球の109倍 | 太陽の2154倍 | 太陽系の3.5万倍 | 天の川銀河の5000倍 |
大きさは円形の直径、数字は全て(約)、1光年=9兆4607億km≒10兆km=1013km
大きさの比率【太陽:地球】≒【直径1m:ビー玉】≒【東京ドーム:直径1m】
1万=104,1億=108,1兆=1012,1京=1016,1垓=1020,太陽系の大きさはオールトの雲の両端(太陽の重力圏)
誕生時の月は地球から2万kmの近くと推定され、潮汐力により公転速度が加速して、46億年後は38万kmに移動
地球から月までの距離は、38万4400Km(光速で片道1.28秒)、月は地球から毎年3.8cmずつ遠ざかっています
地球から太陽までの距離は、1億4960万km(光速で片道8分19秒、太陽は月の400倍も遠い)
天の川銀河のダークマター=光る物質(星やガス)の10倍くらいのダークマター、100万光年
大きさの比率【太陽:地球】≒【直径1m:ビー玉】≒【東京ドーム:直径1m】
1万=104,1億=108,1兆=1012,1京=1016,1垓=1020,太陽系の大きさはオールトの雲の両端(太陽の重力圏)
誕生時の月は地球から2万kmの近くと推定され、潮汐力により公転速度が加速して、46億年後は38万kmに移動
地球から月までの距離は、38万4400Km(光速で片道1.28秒)、月は地球から毎年3.8cmずつ遠ざかっています
地球から太陽までの距離は、1億4960万km(光速で片道8分19秒、太陽は月の400倍も遠い)
天の川銀河のダークマター=光る物質(星やガス)の10倍くらいのダークマター、100万光年
アンドロメダ銀河→局所銀河群→おとめ座銀河団→超銀河団→局所超銀河団→宇宙の大規模構造
銀河群=規模の小さい銀河団、銀河団は銀河の100倍前後、宇宙の大規模構造の大きさは不明
アンドロメダ銀河=天の川銀河の2~2.5倍、地球から253.7万光年、40億年後には天の川銀河と衝突?
銀河群=規模の小さい銀河団、銀河団は銀河の100倍前後、宇宙の大規模構造の大きさは不明
アンドロメダ銀河=天の川銀河の2~2.5倍、地球から253.7万光年、40億年後には天の川銀河と衝突?
大規模構造(泡構造) :超銀河団同士が相互につながって形成する構造
グレートアトラクター:局所超銀河団の銀河を引き寄せている巨大な高密度領域、巨大引力源、銀河の大集団
[グレートアトラクター]=[宇宙のメガロポリス]
[ボイド]=[過疎の村]
グレートウォール :超銀河団は平面状の壁のような分布、銀河フィラメントとも呼ばれる
グレートアトラクター:局所超銀河団の銀河を引き寄せている巨大な高密度領域、巨大引力源、銀河の大集団
[グレートアトラクター]=[宇宙のメガロポリス]
[ボイド]=[過疎の村]
グレートウォール :超銀河団は平面状の壁のような分布、銀河フィラメントとも呼ばれる
| 宇宙空間の地球進行 | 秒速 | 時速 |
|---|---|---|
| 地球の自転 | 460m | 1,700km |
| 地球の太陽系公転 | 30km | 10万km |
| 太陽系の銀河系軌道 | 230km | 83万km |
| 銀河の宇宙空間疾走 | 600km | 216万km |
太陽系は、銀河核中心から約3万光年の軌道を単独で回っています
一周に要する期間、銀河年はおよそ2億2000万年から2億5000万年です
その形成以来、太陽系は銀河系を少なくとも20周したことになります
一周に要する期間、銀河年はおよそ2億2000万年から2億5000万年です
その形成以来、太陽系は銀河系を少なくとも20周したことになります
| 時刻 | 宇宙の年表 | |
|---|---|---|
| 0秒~10-44秒後 | プランク時代 | 超々高温の超々微小宇宙 |
| 10-44秒~10-36秒後 | 大統一時代 | 宇宙が膨張し始め、力はこの時代以降順次分離 |
| 10-36秒~10-34秒後 | インフレーション時代 | 宇宙がものすごい勢いで膨らむ(1030倍以上) |
| 10-34秒~10-11秒後 | 電弱時代(ビッグバン) | 素粒子であふれかえる超高温の火の玉宇宙~超微小宇宙 |
| 10-12秒~10-6秒後 | クォークの時代 | ヒッグス粒子が質量に動きにくさを与える |
| 10-6秒~1秒 | ハドロンの時代:核子 | クォークが合体してできたハドロンは少し残る |
| 1秒~10秒後 | レプトンの時代:電子 | 少しの電子と自由に飛び回る光子とニュートリノ |
| 10秒~38万年後 | 光子時代:光子 | 光子は荷電した陽子、電子、原子核と干渉し38万年続く |
| 3分~20分後 | 原子核の合成 | 陽子と中性子が原子核を作り出す(ビッグバン元素合成) |
| 20分~10万年後 | 原子の形成 | 原子核が電子を捕まえて原子ができる |
| 10万年~38万年後 | 宇宙の晴れ上がり | 自由電子が原子核に束縛され光子が直進(3000度程度) |
| 38万年~数億年後 | 宇宙の暗黒時代 | 漂う原子や自由な光子・ニュートリノだけの真暗宇宙 |
| 2億年~30億年後 | 恒星の誕生~銀河の誕生 | 重力で集まったガス雲→星→銀河→銀河団の形成 |
| 92億年~138億年後 | 地球の誕生~現在の宇宙 | 原始太陽の周りに惑星ができて地球ができる |
プランク時間:光子が光速でプランク長を移動するのにかかる時間=(プランク長÷光速)=5.391×10-44秒
プランク時代 … プランク時間の瞬間は重力は他の力と同じくらい強く、全ての力は統一されていた
大統一時代 … 4つの力のうち重力を除く3つの力(電磁力、強い力、弱い力)が統一されていた
| 時刻 | 予測 |
|---|---|
| 9億年~10億年後 | 太陽光度の上昇による地球生命圏の最後、植物の光合成停止→生物絶滅 植物絶滅のシナリオ:二酸化炭素濃度の減少 3億年後:100ppm、C3植物の光合成が停止 9億年後: 10ppm、C4植物の光合成が停止、ほとんどの生物が絶滅 |
| 15億年~20億年後 | 太陽光度の上昇により地球がハビタブルゾーン(生命生存可能領域)外へ |
| 40億年~60億年後 | 銀河の衝突 → 巨大/楕円銀河に |
| 50億年~75億年後 | 老化する太陽、地球が太陽に飲み込まれるか、急激に蒸発していく(地球の終わり) |
| 1000億年後 | 銀河の孤立(銀河の合体、局所銀河群の合体) |
| 1013年~1015年後 | 恒星が核融合を停止し燃え尽きる → 暗い宇宙 |
| 1014年~1040年後 | 縮退の時代(星の形成の終結) |
| 1019年~1020年後 | 星の残骸物の消滅 |
| 1030年~1033年後 | 銀河・銀河団がすべて中心ブラックホールに飲み込まれる |
| 1034年~1040年後 | 陽子崩壊 → 陽電子・光子・ニュートリノなどに崩壊して物質が消滅 |
| 10100年~10131年後 | ブラックホールの蒸発 → 暗黒の時代 |
生命誕生から40億年後の現在の地球は9億年後が生命圏の最後ならば、老年期に入ったことになる
ホーキング放射の理論
エネルギーから粒子・反粒子が発生する対生成が事象の地平線
近くで生じた場合、正のエネルギーの粒子が外へのがれ、
負のエネルギーの反粒子が落ち、正のエネルギーを失い、
ブラックホールは徐々に消滅に向かいブラックホールが蒸発する
近くで生じた場合、正のエネルギーの粒子が外へのがれ、
負のエネルギーの反粒子が落ち、正のエネルギーを失い、
ブラックホールは徐々に消滅に向かいブラックホールが蒸発する
| 太陽質量の | 恒星の誕生と死 |
|---|---|
| 0.08倍以下 | 星間ガス → 褐色矮星 |
| 0.08倍~8倍 | 星間ガス → 主系列星 → 赤色巨星 → 惑星状星雲 → 白色矮星 |
| 8倍~30倍 | 星間ガス → 主系列星 → 赤色巨星 → 超新星爆発 → 中性子星 |
| 30倍以上 | 星間ガス → 主系列星 → 赤色巨星 → 超新星爆発 → ブラックホール |
星間ガス→主系列星:水素中心核核融合→赤色巨星:ヘリウム中心核・外層水素の核融合
→ヘリウム中心核核融合→炭素や酸素の核・外層ヘリウム核融合→核融合終了→白色矮星
太陽の寿命:100億年(現在46億年経過、残り約50億年)
→ヘリウム中心核核融合→炭素や酸素の核・外層ヘリウム核融合→核融合終了→白色矮星
太陽の寿命:100億年(現在46億年経過、残り約50億年)
ビッグフリーズ(寿命死):宇宙は物質が希薄過ぎて何もないような死の空間になる宇宙の低温死
ビッグクランチ(過収縮):宇宙の膨張が止まり収縮に向かい、最後には宇宙は潰れてしまう
ビッグリップ (過膨張):宇宙の膨張で時空が引き裂かれ、それ以上は宇宙が存続できなくなる
宇宙の熱的死 (熱的死):熱的平衡状態になり、何も起こらなくなる熱力学的な死亡状態
真空の崩壊 (突然死):真空の相転移のような予測不可能な突発的な変化
ビッグバウンス(周期的):特異点で跳ね返り、収縮と膨張を何度も繰り返す
ビッグクランチ(過収縮):宇宙の膨張が止まり収縮に向かい、最後には宇宙は潰れてしまう
ビッグリップ (過膨張):宇宙の膨張で時空が引き裂かれ、それ以上は宇宙が存続できなくなる
宇宙の熱的死 (熱的死):熱的平衡状態になり、何も起こらなくなる熱力学的な死亡状態
真空の崩壊 (突然死):真空の相転移のような予測不可能な突発的な変化
ビッグバウンス(周期的):特異点で跳ね返り、収縮と膨張を何度も繰り返す
| リスク | 理由 | |
|---|---|---|
| 感染症の世界的流行 (パンデミック) | 人間の自然破壊や自然界介入による危険な人獣共通感染症の人への伝染 抗微生物薬耐性の細菌や未知のウイルスの蔓延 人工的な病原体の生成と拡散 | |
| 食料や水資源の枯渇 | 異常気象、人口爆発、経済発展による食料や水需要の激増 天然資源の大量消費による人為的気候変動や生態系の崩壊 | |
| 地球規模の自然現象 | 壊滅的な自然的気候変動による生態系の崩壊や自然災害の増加 | |
| 氷期の到来 | 生物の絶滅や生態系の破壊 | |
| 海洋無酸素イベント | ||
| 洪水玄武岩を噴出する超巨大噴火 | 塵に覆われ寒冷化 | |
| 小惑星の衝突 | ||
| 世界戦争(核戦争) | 大量の核弾頭保有、偶発的 | |
| 世界規模のテロリズム | 貧困の拡大、宗教や思想の対立、資源の争奪、暴力的過激思想の拡散 貧者の兵器による戦争の拡大(生物化学兵器,小型核兵器,自爆ドローン) | |
| 人工知能やロボットの暴走 | 自立兵器の乱用や粗悪な開発 制御不能な誤動作や反乱 | |
| ナノテクの誤用 | 自己増殖能力を持つナノマシン | |
| 大規模なシステム崩壊 | 世界規模での経済システムや社会システムの崩壊 一部に起こる想定外事態や人的失敗による全体システムの破綻 | |
|
英オックスフォード大とスウェーデンの財団等で構成する研究チームが2015年発表 「12 Risks That Threaten Human Civilization」(人類の文明を脅かす12のリスク) Current risks 現在のリスク 1. Extreme Climate Change 極端な気候変動 2. Nuclear War 核戦争 3. Ecological Catastrophe 生態系の崩壊 4. Global Pandemic 世界規模のパンデミック 5. Global System Collapse 国際的な経済・社会システムの崩壊 Exogenic risks 外因性リスク 6. Major Asteroid Impact 巨大隕石の衝突 7. Supervolcano 大規模な火山噴火 Emerging risks 新たなリスク 8. Synthetic Biology 合成生物学(病原体の生成) 9. Nanotechnology ナノテクノロジー 10. Artificial Intelligence 人工知能 11. Uncertain Risks 未知の可能性 Global policy risks グローバルポリシーリスク 12. Future Bad Global Governance 劣悪なグローバル・ガバナンス |
「人類を不妊にする超汚染物質の開発」
「人工ブラックホールが開発され、地球を飲み込むこと」
「動物実験により人類を超える知能をもつ生物が出現」
「誰かが地球外生命にコンタクトし、危険な異星人の注意を呼び寄せること」
14世紀:黒死病(ペスト)、1520年:天然痘、19~20世紀:コレラ、1918~1920年:スペインかぜ、
1981年:エイズ(HIV)、2002年:重症急性呼吸器症候群(SARS)、2009年:新型インフルエンザ(A/H1N1)、
2020年3月11日:WHOが新型コロナウイルス(COVID-19)のパンデミック宣言
1981年:エイズ(HIV)、2002年:重症急性呼吸器症候群(SARS)、2009年:新型インフルエンザ(A/H1N1)、
2020年3月11日:WHOが新型コロナウイルス(COVID-19)のパンデミック宣言
潜在的に危険な小惑星(PHA):1300個以上発見、全PHAの20~30%、ほとんどは未発見、隕石落下:年平均40回程度
今後100年以内では878個の小惑星が地球に衝突するリスクがあるようです(PHA:Potentially Hazardous Asteroid)
恐竜を絶滅に追いこんだ小惑星の直径は約10kmです … メキシコのユカタン半島海底のクレーターは直径180km
1994年SL第9彗星が木星重力に引き寄せられ、潮汐力で直径1~5kmに分裂した破片が秒速60kmで木星に衝突した
時の衝撃は広島に投下された原爆の1億倍~10億倍と概算され、衝突時のきのこ雲が地上の望遠鏡でも観測された
2013年ロシアのチェリャビンスク州で大きな隕石が落下、NASAの推定によれば、直径17m、重量七千トン、地球へ
の入射速度は18km/s、エネルギーは広島原爆30個分、損害は死者0人、負傷者1491人、4474棟の建造物が損壊した
今後100年以内では878個の小惑星が地球に衝突するリスクがあるようです(PHA:Potentially Hazardous Asteroid)
恐竜を絶滅に追いこんだ小惑星の直径は約10kmです … メキシコのユカタン半島海底のクレーターは直径180km
1994年SL第9彗星が木星重力に引き寄せられ、潮汐力で直径1~5kmに分裂した破片が秒速60kmで木星に衝突した
時の衝撃は広島に投下された原爆の1億倍~10億倍と概算され、衝突時のきのこ雲が地上の望遠鏡でも観測された
2013年ロシアのチェリャビンスク州で大きな隕石が落下、NASAの推定によれば、直径17m、重量七千トン、地球へ
の入射速度は18km/s、エネルギーは広島原爆30個分、損害は死者0人、負傷者1491人、4474棟の建造物が損壊した
直径1kmほどの小惑星が衝突しても、全人口の10%は失われると試算されています
日本のJAXAやアメリカのNASAで打ち上げられた小惑星探査機の使命は「宇宙・太陽系の科学的探査」だけではなく、
もう一つの「将来の潜在的に危険な小惑星(PHA)に人類が対処する方法を探る」という重要な使命を帯びています
欧州宇宙機関やNASAは、宇宙機を小惑星に衝突させて地球に向かう小惑星の軌道を変える地球防衛の実験を計画
NASAは2022年09月26日無人探査機DARTを小惑星ディモルフォスに体当たりさせる「惑星防衛」の実証実験を行った
日本のJAXAやアメリカのNASAで打ち上げられた小惑星探査機の使命は「宇宙・太陽系の科学的探査」だけではなく、
もう一つの「将来の潜在的に危険な小惑星(PHA)に人類が対処する方法を探る」という重要な使命を帯びています
欧州宇宙機関やNASAは、宇宙機を小惑星に衝突させて地球に向かう小惑星の軌道を変える地球防衛の実験を計画
NASAは2022年09月26日無人探査機DARTを小惑星ディモルフォスに体当たりさせる「惑星防衛」の実証実験を行った
2022.01.03:国連安保理の常任理事国(核保有国5か国米中露英仏)は、「核兵器国の間の戦争回避が最も重要な責務」
「核戦争に勝者はなく、決して戦ってはならないことを確認する」と声明した
原爆死没者慰霊碑の碑文「安らかに眠ってください 過ちは繰り返しませぬから」
Let all the souls here rest in peace ; For we shall not repeat the evil
「核戦争に勝者はなく、決して戦ってはならないことを確認する」と声明した
原爆死没者慰霊碑の碑文「安らかに眠ってください 過ちは繰り返しませぬから」
Let all the souls here rest in peace ; For we shall not repeat the evil
平等は破壊の後にやってくる → 不平等を是正してきたのは、「戦争・革命・崩壊・疫病」
格差による感染症のリスクと格差の拡大、貧しい国のテロリストが裕福な国に移動、他国への移住が止められない
世界経済がグローバル化して、経済危機や貧富の差が拡大し、「格差の世襲化」は社会混乱や無法状態をもたらす
現在の先進国は端的にいえば、「持続可能性がないコース」を突き進んでいます … 文明の崩壊リスク
格差による感染症のリスクと格差の拡大、貧しい国のテロリストが裕福な国に移動、他国への移住が止められない
世界経済がグローバル化して、経済危機や貧富の差が拡大し、「格差の世襲化」は社会混乱や無法状態をもたらす
現在の先進国は端的にいえば、「持続可能性がないコース」を突き進んでいます … 文明の崩壊リスク
▲ YouTube
核戦争の恐怖 → ターミネーター2 サラ・コナーの悪夢(核爆発の炎が押し寄せる)
▲ YouTube
太陽フレア → ノウイングのラストシーン(地球上が建物ごと炎の波に流される)
▲ YouTube
核戦争後 → テイラーが自由の女神像を見つける(猿の惑星は地球だった)
核戦争の恐怖 → ターミネーター2 サラ・コナーの悪夢(核爆発の炎が押し寄せる)
▲ YouTube
太陽フレア → ノウイングのラストシーン(地球上が建物ごと炎の波に流される)
▲ YouTube
核戦争後 → テイラーが自由の女神像を見つける(猿の惑星は地球だった)
|
未来社会構想 2050「豊かで持続可能な世界」を実現するために デジタル経済圏の台頭 2050年にかけて、プラットフォーマーなどが発行するデジタル通貨や、それにひもづく経済活動 が拡大し、デジタル経済圏が形成される 覇権国のいない国際秩序 国際社会では絶対的な覇権国のいない世界が実現する 「デジタル移民」の増加と広がる国内格差、高度人材比率が国の成長の鍵 脱炭素を実現する循環型社会 デジタル技術のさらなる普及は、循環型社会の実現を後押しする 太陽光や風力など再生可能エネルギーを軸とした需給構造、資源面はリサイクル・代替が加速する 変容する政府の役割 行政サービスの効率化、国際的なルールの策定や順守体制の構築、デジタル経済圏の拡大に伴う 新しい環境整備、経済格差に対するセーフティーネットの提供などで、政府の役割が拡大しよう 多様なコミュニティが共存する社会 デジタル空間中で議論や主張をする際、情報の偏りや意見の偏りのようなコミュニティの分断が 深まる恐れがある 技術によって変わる人生 さまざまなイノベーションの実装に伴って、経済活動の半分以上はデジタル経済圏に関わるものに なり、家事の自動化や通勤時間が減ることで自由時間が増え、健康寿命も延伸される 実現に向けて必要なアクション(2050年の日本) 「人間中心の技術活用」データ・AI・ロボット 「日本の良さ・強み」和・匠・美、「前向きな挑戦」で社会を変革 |
いくつかの国の、力による領土拡張や一方的現状変更は、当事国や近隣諸国との軋轢を生み、戦争の危険性を孕んでいます
| SDGsの17の目標 |
|---|
|
SDGsは、2015年9月の国連サミットで全会一致で採択された「持続可能な開発のための2030アジェンダ」に掲げられ た、「持続可能な開発目標(Sustainable Development Goals)」のことです SDGsは「17の目標」と「169のターゲット(具体目標)」で構成されています 5つのP … 人間:People、豊かさ:Prosperity、地球:Planet、平和:Peace、パートナーシップ:Partnership 1~6 7~12 13~15 16 17
┌── 1.貧困をなくそう │ 2.飢餓をゼロに │ 3.すべての人に健康と福祉を ┌─「社会」──┤ 4.質の高い教育をみんなに │ │ 5.ジェンダー平等を実現しよう │ └── 6.安全な水とトイレを世界中に │ ┌── 7.エネルギーをみんなに そしてクリーンに │ │ 8.働きがいも経済成長も SDGsの17目標リスト─┼─「経済」──┤ 9.産業と技術革新の基盤をつくろう │ │ 10.人や国の不平等をなくそう 社会・経済・環境 │ │ 11.住み続けられるまちづくりを の3分野と枠組み │ └──12.つくる責任つかう責任 │ ┌──13.気候変動に具体的な対策を ├─「環境」──┤ 14.海の豊かさを守ろう │ └──15.陸の豊かさも守ろう └─「枠組み」─┬──16.平和と公正をすべての人に └──17.パートナーシップで目標を達成しよう ※ただし、一部のカルト宗教団体は勧誘手段としてSDGsを悪用しているものもありますので要注意です |
| 量子ゆらぎ→インフレーション→密度ゆらぎ→銀河や銀河団の成長 | |
|---|---|
| 無からの宇宙創成 + インフレーション理論 量子宇宙論 | 真空の状態 |
| 無は宇宙の種が量子ゆらぎしている時空の泡状態(粒子・反粒子の生成消滅繰返し) | |
| 宇宙の種→確率的なトンネル効果で超々微小宇宙が誕生する | |
| 重力の分岐 | |
| 宇宙のインフレーション | |
| 超々微小宇宙とともに量子ゆらぎも大きくひきのばされて | |
| 現在の銀河や銀河団の種になる(エネルギー密度のムラ:密度ゆらぎ) | |
| 強い力の分岐 | |
| ビッグバン理論 ↓ 火の玉宇宙→宇宙の膨張 一般相対性理論 | ビッグバン |
| インフレーション終了→潜熱の解放→全エネルギーが熱に変換→物質と光の誕生 | |
| 対称性の自発的破れ(真空の相転移) | |
| 冷たい宇宙──────→熱い宇宙 | |
| 真空のエネルギーが物質と光に転化 | |
| 弱い力・電磁気力の分岐 | |
| 物質の誕生(素粒子→核子→原子核→原子) | |
| ヒッグス粒子が作用する → 質量に動きにくさを与える → 素粒子の誕生 | |
| 色の力が作用する → クォークの閉じ込め → 核子の誕生 | |
| 核力が作用する → 核子の結合(核融合) → 原子核の合成 | |
| 電気の力が作用する → 電子が原子核に束縛される → 原子の形成 | |
| 宇宙の大規模構造形成 | |
| 重力が作用する → 原子が集まる(星間ガス雲) → 恒星の誕生 | |
| 星・銀河・銀河団の形成、超銀河団の連なりと超空洞が網の目構造を形成 | |
| 太陽系の誕生、地球・月の誕生 | |
| 生命誕生、人類誕生 | |
量子の世界において、電子が古典的には超えられないエネルギー障壁を一部反射せず通り抜ける現象
質量が大きいほど、トンネル効果は起きにくくなる … 不確定性原理⊿x⊿p≧h/4π
(x:位置=波の広がり、p:運動量=質量×速度)
量子トンネリングは障壁の厚さがおよそ1–3nm以下の場合に起こる(1nm=10億分の1メートル)
ヒッグス機構:宇宙創成の直後は真空(ヒッグス場)は水蒸気のように透明に広がっている状態ですべての
素粒子は質量がなく光速で飛びまわったいたのですが、対称性の破れが起こりエネルギー状態が変わると
水蒸気が水滴に変わるようにヒッグス粒子は粒子として出現しこれがくっついて素粒子は動きにくくなり
質量が誕生しました
| 宇宙創成後の宇宙の姿を変えた謎の解明 | 新理論 |
|---|---|
| 対称性の自発的破れの理論 (CP対称性の破れなど) | 真空の相転移=対称性の自発的破れ |
| インフレーション宇宙モデル | |
| ヒッグス粒子による質量の誕生(ヒッグス機構) | |
| 消えた反粒子(CP対称性の破れのおかげ)← 未解明 |
元の物質を構成する素粒子のすべてにC変換を施すと反物質になる
CP対称性の破れ … クォークの変身の種類が増すことにより破れが生じる可能性がある
そのことによりその分の反物質が消滅する
宇宙の物理法則 … 対称性は何種類もあり、どの対称性がどの程度破れているかによって、
宇宙の物理法則が決まっている
万有引力定数やクーロン定数がいまある値になったのも、
対称性の破れ具合によるものです
| 未解明の謎 | ||
|---|---|---|
| 宇宙の歴史 | 宇宙創成の瞬間 | 真空の相転移とは、真空の基底状態が変化することか |
| 消えた反物質 | バリオン数生成問題、物質・反物質の対生成・対消滅の差 | |
| 暗黒物質 | 宇宙の見えない質量、ゆっくり動く質量の大きな粒子 | |
| 暗黒エネルギー | 宇宙の加速膨張、負の圧力(重力斥力)を持つ仮想的なエネルギー | |
| ブラックホール | 光さえも脱出できないほど重力が強い天体 | |
| 宇宙の果て | 果ての有無、観測可能性 | |
| 人類の未来 | 人類の滅亡リスク | パンデミック・小惑星の衝突 |
| 宇宙創成理論 | 量子宇宙論 | 量子論に基づいて宇宙の始まりを考える |
| 真空のエネルギー | 時空の泡状態(粒子・反粒子の生成消滅繰返し)、ゼロ点エネルギー | |
| 重力波 | 光速で伝わる時空のさざ波、超新星爆発などが発生源 | |
| 地質時代 | 生命の起源 | 宇宙起源説・地球起源説 |
| 地球外生命 | 地球外に生命を探す宇宙探査 | |
| 未来の人類 | 人類の進化の方向、超人?、機械との融合? | |
| 地球史年表 | 人新世 | 人類が地球の生態系や気候に大きな影響を及ぼすようになった現代 |
| 生物系統図 | 起源生物 | 全生物共通祖先(コモノート) |
| ミクロの世界 | 大統一理論 | 強い力と電弱力を統合 |
| ミクロな粒子 | ニュートリノ振動 | 宇宙線起源の大気ニュートリノの変化 |
| 超対称性理論 | フェルミ粒子とボース粒子の組が対称な超対称性粒子を考える理論 | |
| 陽子崩壊 | クォークからなる陽子がレプトン族に変換され、物質が消滅する | |
| 超弦理論 | 粒子でなくひも、ひもの振動の強弱で素粒子の種類が決まる | |
| ヒッグス粒子 | ヒッグス粒子による質量の誕生(ヒッグス機構) | |
| 量子論 | 多世界解釈 | 多数の世界の同時存在 |
提案1:反物質の寿命がわずかに短かったので、反物質だけが消滅し、物質だけが取り残された
提案2:物質が反物質よりも少しだけ余計にあって、対消滅を免れた
提案3:粒子と反粒子の反応法則に、ごくわずかの違いがあり、その差の分だけ粒子が残った
提案4:ニュートリノと反ニュートリノの同一性を実証すれば、反粒子が粒子に変わり得ること
がわかり、粒子と反粒子の数量の比が変化することが説明できる
レプトジェネシス機構
宇宙初期の相転移が、ニュートリノに物質と反物質の入れ替えを可能とさせたとする理論
ニュートリノは電気的に中性な粒子であり、反物質から物質の入れ替えが可能な最も有力
な候補とされています
対消滅をまぬがれた素粒子は10億個のうちわずか2個程度だった
バリオン数生成問題
初期宇宙のインフレーションによって、仮にそれ以前に正味バリオン数があったとしても
薄まってしまうので、観測されているバリオン非対称は、すべてインフレーション後に作
られなければならない
物質・反物質対生成の偏りが生じたイベントのことを「バリオン数生成」と呼んでいる
実証済みの事柄
対消滅:電子と陽電子が衝突してエネルギーとなり物質は消滅(対消滅)して光になる
対生成:空間の1点にエネルギーが集中すると、粒子と反粒子が誕生する(対生成)
①高エネルギー光子同士の衝突
光子 + 光子 → 電子 + 陽電子
光のエネルギーが相対性理論の効果で物質に変わるからです
②更にエネルギーを上げると質量の大きい粒子や多数の粒子が一度に作られるようになる
レプトン(電子)ができれば、バリオン(陽子、中性子)も作れる
電子 + 陽電子 → 陽子 + 反陽子
①高いエネルギーの光子同士の衝突
光と光がぶつかっても消えてなくなったり、跳ね返ったり、地面に落ちたりはしません
素通りするだけです
しかし、エネルギーの高い光である「ガンマ線」同士をぶつけると、反応が起こります
これは、光のエネルギーが相対性理論の効果で物質に変わるからです
光子1個のエネルギーが、電子の質量に対応するエネルギー(0.511MeV)より大きくなると、
2個の光子の衝突により、電子とその反粒子である陽電子のペアが作られます
2個の光子は消えてなくなってしまいます … 光子 + 光子 → 電子 + 陽電子
②更にエネルギーを上げると質量の大きい粒子や多数の粒子が一度に作られるようになります
重い中間子の生成(0.5~5GeV程度の光子で起こる)や
エネルギーの高いクォークやグルーオンから作られる多数の粒子群(ハドロン)の生成
(おもに2GeV以上の光子による)
※1MeV=100万電子ボルト、1GeV=10億電子ボルト
提案2:物質が反物質よりも少しだけ余計にあって、対消滅を免れた
提案3:粒子と反粒子の反応法則に、ごくわずかの違いがあり、その差の分だけ粒子が残った
提案4:ニュートリノと反ニュートリノの同一性を実証すれば、反粒子が粒子に変わり得ること
がわかり、粒子と反粒子の数量の比が変化することが説明できる
レプトジェネシス機構
宇宙初期の相転移が、ニュートリノに物質と反物質の入れ替えを可能とさせたとする理論
ニュートリノは電気的に中性な粒子であり、反物質から物質の入れ替えが可能な最も有力
な候補とされています
粒子 反粒子
○○○○……○○○○ ●●●●……●●●●
○(10億+1)個 ●(10億+1)個
○○○○……○○○○ ●●●●……●●●●
○(10億+1)個 ●(10億+1)個
粒子 反粒子
○○○○……○○○○ ●●●●……●●●●
○○(10億+2)個 ○←──● (10億)個
○○○○……○○○○ ●●●●……●●●●
○○(10億+2)個 ○←──● (10億)個
反粒子の1個が普通の粒子に
粒子 反粒子
10億のペアが対消滅
○○2個 0個
10億のペアが対消滅
○○2個 0個
バリオン数生成問題
初期宇宙のインフレーションによって、仮にそれ以前に正味バリオン数があったとしても
薄まってしまうので、観測されているバリオン非対称は、すべてインフレーション後に作
られなければならない
物質・反物質対生成の偏りが生じたイベントのことを「バリオン数生成」と呼んでいる
実証済みの事柄
対消滅:電子と陽電子が衝突してエネルギーとなり物質は消滅(対消滅)して光になる
対生成:空間の1点にエネルギーが集中すると、粒子と反粒子が誕生する(対生成)
①高エネルギー光子同士の衝突
光子 + 光子 → 電子 + 陽電子
光のエネルギーが相対性理論の効果で物質に変わるからです
②更にエネルギーを上げると質量の大きい粒子や多数の粒子が一度に作られるようになる
レプトン(電子)ができれば、バリオン(陽子、中性子)も作れる
電子 + 陽電子 → 陽子 + 反陽子
①高いエネルギーの光子同士の衝突
光と光がぶつかっても消えてなくなったり、跳ね返ったり、地面に落ちたりはしません
素通りするだけです
しかし、エネルギーの高い光である「ガンマ線」同士をぶつけると、反応が起こります
これは、光のエネルギーが相対性理論の効果で物質に変わるからです
光子1個のエネルギーが、電子の質量に対応するエネルギー(0.511MeV)より大きくなると、
2個の光子の衝突により、電子とその反粒子である陽電子のペアが作られます
2個の光子は消えてなくなってしまいます … 光子 + 光子 → 電子 + 陽電子
②更にエネルギーを上げると質量の大きい粒子や多数の粒子が一度に作られるようになります
重い中間子の生成(0.5~5GeV程度の光子で起こる)や
エネルギーの高いクォークやグルーオンから作られる多数の粒子群(ハドロン)の生成
(おもに2GeV以上の光子による)
※1MeV=100万電子ボルト、1GeV=10億電子ボルト
オパーリンが唱えた物質の化学進化説
還元的環境を前提としたユーリー・ミラーの実験
(予想した原始地球の大気組成に高電圧火花放電=雷を発生させ1週間後にアミノ酸生成)
は、地球における有機物の誕生を再現したものとは言えないことになった
一次大気(原始大気):還元的組成、水素、メタン、アンモニア ユーリーミラーの実験
二次大気(初期大気):酸化的組成、二酸化炭素、窒素 当時の地球大気
新たな有機物生成過程
化学進化の第一段階である有機物合成には、
当時の地球大気を再現していないユーリー・ミラーの実験に代わる、別の過程が必要になる
その後、マグマオーシャン中には金属鉄が含まれ、大気と金属鉄の反応によって初期大気は
二次大気組成だけでなく水素や一酸化炭素を大量に含む還元的組成になることが判明した
パンスペルミア説が「宇宙起源説」、他は「地球起源説」をとる
表面代謝説 (粘土界面上のアミノ酸重合反応)
パンスペルミア説 (生命のタネは宇宙の星間分子雲から飛来した)
深海熱水孔での独立栄養生物説 (最初の生命は独立栄養生物だった)
分子生物学的化学進化説 (地球から細胞が生まれた)
還元的環境を前提としたユーリー・ミラーの実験
(予想した原始地球の大気組成に高電圧火花放電=雷を発生させ1週間後にアミノ酸生成)
は、地球における有機物の誕生を再現したものとは言えないことになった
一次大気(原始大気):還元的組成、水素、メタン、アンモニア ユーリーミラーの実験
二次大気(初期大気):酸化的組成、二酸化炭素、窒素 当時の地球大気
新たな有機物生成過程
化学進化の第一段階である有機物合成には、
当時の地球大気を再現していないユーリー・ミラーの実験に代わる、別の過程が必要になる
その後、マグマオーシャン中には金属鉄が含まれ、大気と金属鉄の反応によって初期大気は
二次大気組成だけでなく水素や一酸化炭素を大量に含む還元的組成になることが判明した
パンスペルミア説が「宇宙起源説」、他は「地球起源説」をとる
表面代謝説 (粘土界面上のアミノ酸重合反応)
パンスペルミア説 (生命のタネは宇宙の星間分子雲から飛来した)
深海熱水孔での独立栄養生物説 (最初の生命は独立栄養生物だった)
分子生物学的化学進化説 (地球から細胞が生まれた)
宇宙では、重い元素からなるダストに宇宙線や紫外線が当たり、有機物ができる
隕石が落下して地球に衝突し、生命の材料(有機物)がたどりつき、エネルギーが供給され、
深海の熱水噴出孔で金属イオンを触媒にして化学的な進化をとげた
隕石が落下して地球に衝突し、生命の材料(有機物)がたどりつき、エネルギーが供給され、
深海の熱水噴出孔で金属イオンを触媒にして化学的な進化をとげた
隕石中に生命の源か
炭素質コンドライト隕石(マーチソン隕石)には右手型と左手型のアミノ酸が含まれていた
地球上の生物はタンパク質生成の際に左手型だけを使う
地球上の生物はタンパク質生成の際に左手型だけを使う
深海の熱水噴出口が生命誕生の場所
原料分子(H2O,CO2,CO,N2)→低分子素材→高分子部品(タンパク質,糖類,核酸)→組立→小胞
以下はどの説でも大枠は同じ
アミノ酸をペプチド結合させ(アミノ酸の重合)タンパク質(高分子化合物)を合成、
糖質や脂質、核酸も合成された
糖質はエネルギー源のグルコース、脂質は細胞膜、核酸はDNAやRNAとなり、小胞が形成され、
細胞の原型ができた(DNA:デオキシリボ核酸、RNA:リボ核酸)
細胞の基本機能は進化し、代謝や細胞形状の形成や自己複製機能(遺伝子)を持つに至った
共通祖先→原核生物(真正細菌、古細菌)、真核生物(細胞内にさまざまな細胞内小器官をもつ)
原料分子(H2O,CO2,CO,N2)→低分子素材→高分子部品(タンパク質,糖類,核酸)→組立→小胞
以下はどの説でも大枠は同じ
アミノ酸をペプチド結合させ(アミノ酸の重合)タンパク質(高分子化合物)を合成、
糖質や脂質、核酸も合成された
糖質はエネルギー源のグルコース、脂質は細胞膜、核酸はDNAやRNAとなり、小胞が形成され、
細胞の原型ができた(DNA:デオキシリボ核酸、RNA:リボ核酸)
細胞の基本機能は進化し、代謝や細胞形状の形成や自己複製機能(遺伝子)を持つに至った
共通祖先→原核生物(真正細菌、古細菌)、真核生物(細胞内にさまざまな細胞内小器官をもつ)
DNAが遺伝情報保存、RNAを仲介として、タンパク質を発現する流れ(セントラルドグマ)では、
DNAをつくる際にはDNA合成酵素、DNAからRNAをつくる際にはRNA合成酵素が必要です
DNA合成酵素もRNA合成酵素もタンパク質です
タンパク質を作るためにはDNAが必要ですが、
DNAからタンパク質を作る際には、反応を触媒する酵素(タンパク質)が必要です
化学進化後の最初の生命でこれら3つの物質のいずれが雛形となったのか、以下の諸説ある
DNAワールド仮説:DNAの情報をコピーしたRNAをもとにタンパク質をつくるので、DNAが先
RNAワールド仮説:RNAが遺伝情報と触媒機能の両方を担う
太古の昔はRNAが遺伝子を作っていたと考えられています
太古の生命体はDNAやタンパク質を使わずにRNAだけでできており
それが情報と機能の両方を兼ね備えていたという説
RNAがタンパク質と同じような触媒機能をもつ(酵素としてはたらく)という発見だった
遺伝子として情報を保持する働きも、実際に活動するタンパク質としての働きもRNAが
すべて担っていた ← 触媒としてはたらくRNA(リボ核酸)や逆転写酵素の発見
RNAワールド仮説の下では、まず初めに、自己複製機能と様々な反応の触媒機能を持つ
(つまり、代謝も担うことができる)RNA分子(リボザイム)が出現したと考える
それらのRNA分子が「進化」の結果複雑化していき、
RNAの代わりに代謝を担うタンパク質を生成するようになったと考えられている
RNAの情報をタンパク質へと読み換えるプロセスである「翻訳(タンパク質合成)」の
メカニズムの発生が生命の「進化」に必須であったということになる
当初はRNAだけで遺伝も触媒もになっていたものが、
触媒に効率のいいタンパク質を使うようになった
一方、遺伝情報の量が増加し複雑になると、RNAより安定なDNA、
それもより安定な二本鎖DNAに情報を保存するようになり、
遺伝子を貯蔵するにも安定したDNAを使い始める
DNAからRNAへの「転写(必要部分のコピー)」が始まったと考えられています
またこのように一段階増やすことによって、より複雑な調節が可能になりました
DNAをつくる際にはDNA合成酵素、DNAからRNAをつくる際にはRNA合成酵素が必要です
DNA合成酵素もRNA合成酵素もタンパク質です
タンパク質を作るためにはDNAが必要ですが、
DNAからタンパク質を作る際には、反応を触媒する酵素(タンパク質)が必要です
化学進化後の最初の生命でこれら3つの物質のいずれが雛形となったのか、以下の諸説ある
DNAワールド仮説:DNAの情報をコピーしたRNAをもとにタンパク質をつくるので、DNAが先
複製 転写 翻訳
DNA─→DNA─→RNA─→タンパク質
逆転写 (アミノ酸)
DNA←─RNA
プロテインワールド仮説:合成酵素はタンパク質なので、タンパク質が先(GADV仮説も出現)DNA─→DNA─→RNA─→タンパク質
逆転写 (アミノ酸)
DNA←─RNA
RNAワールド仮説:RNAが遺伝情報と触媒機能の両方を担う
太古の昔はRNAが遺伝子を作っていたと考えられています
太古の生命体はDNAやタンパク質を使わずにRNAだけでできており
それが情報と機能の両方を兼ね備えていたという説
RNAがタンパク質と同じような触媒機能をもつ(酵素としてはたらく)という発見だった
遺伝子として情報を保持する働きも、実際に活動するタンパク質としての働きもRNAが
すべて担っていた ← 触媒としてはたらくRNA(リボ核酸)や逆転写酵素の発見
RNAワールド仮説の下では、まず初めに、自己複製機能と様々な反応の触媒機能を持つ
(つまり、代謝も担うことができる)RNA分子(リボザイム)が出現したと考える
それらのRNA分子が「進化」の結果複雑化していき、
RNAの代わりに代謝を担うタンパク質を生成するようになったと考えられている
RNAの情報をタンパク質へと読み換えるプロセスである「翻訳(タンパク質合成)」の
メカニズムの発生が生命の「進化」に必須であったということになる
当初はRNAだけで遺伝も触媒もになっていたものが、
触媒に効率のいいタンパク質を使うようになった
一方、遺伝情報の量が増加し複雑になると、RNAより安定なDNA、
それもより安定な二本鎖DNAに情報を保存するようになり、
遺伝子を貯蔵するにも安定したDNAを使い始める
DNAからRNAへの「転写(必要部分のコピー)」が始まったと考えられています
またこのように一段階増やすことによって、より複雑な調節が可能になりました
① RNA
↓ (翻訳)
② RNA─→タンパク質
↓ (転写) (翻訳)
➂ DNA─→RNA─→タンパク質
最初の生命はRNAワールド仮説で説明するのが主流、現在の生命はDNAワールドです↓ (翻訳)
② RNA─→タンパク質
↓ (転写) (翻訳)
➂ DNA─→RNA─→タンパク質
| 地質時代 | 開始年代 | 出来事 | 動物界 | 植物界 | ||
|---|---|---|---|---|---|---|
| 先カンブリア時代 | 冥王代 | 45億6700万年前 | 地球の誕生 | |||
| 45億年前 | 小天体の重爆撃、巨大衝突 (ジャイアントインパクト) により月の誕生 | ダスト表面 → 有機物(アミノ酸等) 糖,脂質,ペプチド,核酸の合成→小胞 | ||||
| 44億年前 | 原始大気と原始海洋の出現 海洋と大陸地殻の形成 (マグマで地表更新→地殻破壊) | |||||
| 41億年前 | 始原生物(共通祖先)の出現 | 原始生命体(代謝系,細胞膜,自己複製) 役割受持ち(蛋白質,糖脂質,核酸DNA) | ||||
| 太古代 | 原太古代(40) 古太古代(36) 中太古代(32) 新太古代(28) | 40億3100万年前 | 陸地の出現(アカスタ片麻岩) 堆積岩中のジルコン粒子包有物 グラファイト(光合成生物の痕跡) | 単細胞生物(40-38)億年前 嫌気性微生物(真正細菌) メタン菌(古細菌) 原核生物:真正細菌、古細菌 微化石:微生物の化石(34)億年前 | ||
| 38億年前 | 小天体後期重爆撃(39-38)億年前 地磁気の形成(38-27)億年前 | |||||
| 27億年前 | 光合成生物の進化(30-25)億年前 | シアノバクテリア(酸素発生型光合成) | ||||
| 原生代 | 古原生代(25) 中原生代(16) 新原生代(10) | 25億年前 | 全球凍結(23)億年前 大酸化イベント(24.5-22)億年前 縞状鉄鉱層の形成(25-19)億年前 | 共生共進化(寄生→相利、任意→必須) 細胞内共生(ミトコンドリア・葉緑体) 真核生物(21)億年前、グリパニア | ||
| 7億年前 | 全球凍結(7)億年前 後期酸化イベント(7)億年前 | 全球凍結下生物痕跡?クリオコナイト (雪氷微生物由来の有機物を含む粒) | ||||
| エディアカラ紀 | 6億3500万年前 | 全球凍結(6.35)億年前 後期酸化イベント(6.35)億年前 エディアカラ生物群(無殻軟組織) | 胚化石 (真正後生動物?) 多細胞生物(6-5.5)億年前 | |||
| 顕生代 | 古生代 | カンブリア紀 | 5億3880万年前 | カンブリア大爆発(硬い組織と眼) | 三葉虫の出現 | 藻類の出現 |
| オルドビス紀 | 4億8540万年前 | 生物大放散事変(生物多様化促進) 植物の陸上進出(コケ植物) | オウムガイの 全盛期 | 藻類の繁栄 | ||
| シルル紀 | 4億4340万年前 | 昆虫の出現(4.8)億年前↑ | 魚類の出現 | シダ植物の出現 | ||
| デボン紀 | 4億1920万年前 | 昆虫の陸上進出 動物の陸上進出、有翅昆虫出現 | 魚類の繁栄 両生類の出現 | シダ類森林(リンボク) 裸子植物の出現 | ||
| 石炭紀 | 3億5890万年前 | 軟骨魚類(サメ)の進化 昆虫類の大繁栄時代(巨大化) | 爬虫類の出現 両生類の繁栄 | 木生シダ植物の繁栄 (大森林時代) | ||
| ペルム紀 | 2億9890万年前 | 超大陸パンゲアの誕生 単弓類の出現と絶滅 | 三葉虫の絶滅 | 木生シダ植物の衰退 (種子植物に交代) | ||
| 中生代 | 三畳紀 | 2億5190万年前 | 最大規模の大量絶滅(PT境界) 哺乳類の祖が出現 | 爬虫類の繁栄 恐竜の出現 | 裸子植物の繁栄 ソテツ、イチョウ 針葉樹(南洋杉など) | |
| ジュラ紀 | 2億0130万年前 | アンモナイトの繁栄、始祖鳥 | 恐竜の繁栄 | |||
| 白亜紀 | 1億4500万年前 | 鳥類の祖(羽毛恐竜)が出現 小惑星の衝突 | 哺乳類の進化 恐竜の絶滅 | 被子植物の出現 (虫媒花の進化) | ||
| 新生代 | 古第三紀 | 6600万年前 | 大型恐鳥類、クジラの祖先 霊長類の出現、飛ぶ鳥の繁栄 | 哺乳類の繁栄 サンゴ礁 | 被子植物の繁栄 熱帯雨林 | |
| 新第三紀 | 2303万年前 | 霊長類の進化(猿類→類人猿) | 人類の出現 | メタセコイア(針葉樹) | ||
| 第四紀 | 258万年前 | 氷河期、土器の発明、農耕牧畜時代 石器時代→青銅器時代→鉄器時代 先史時代→有史時代 | 大型哺乳類の 繁栄と絶滅 人類の繁栄 | ワタスゲ(被子植物) コケ類(ゼニゴケなど) | ||
地質時代は常に見直されており、特に先カンブリア時代の出来事は年代数値も含め異説があり更新されます
最古の木:デボン紀後期の前裸子植物アーケオプテリス(30m)が河川沿いに生息域を拡大し最古の森林を形成
※年代は国際層序委員会,2023年09月によります(ただし三畳紀の開始年代は2億5190万年前に未満省略)
最古の木:デボン紀後期の前裸子植物アーケオプテリス(30m)が河川沿いに生息域を拡大し最古の森林を形成
※年代は国際層序委員会,2023年09月によります(ただし三畳紀の開始年代は2億5190万年前に未満省略)
示準化石:その化石を含む地層が堆積した時代がわかる化石、三葉虫、アンモナイト、貨幣石、コノドント
示相化石:その化石を含む地層が堆積した当時の環境がわかる化石、造礁サンゴ、アサリ、シジミ、タニシ
※体化石:殻・骨格・花粉・胞子など体の全体や一部、生痕化石:生活痕跡、化学化石:生物起源の有機物
示相化石:その化石を含む地層が堆積した当時の環境がわかる化石、造礁サンゴ、アサリ、シジミ、タニシ
※体化石:殻・骨格・花粉・胞子など体の全体や一部、生痕化石:生活痕跡、化学化石:生物起源の有機物
地球の内部構造は花崗岩からなる大陸地殻と玄武岩からなる海洋地殻、マントル、外核、内核
地殻と硬いマントルの最上部をプレート、プレートを動かすマントルの水平対流とプレートの
下降流をコールドプルーム、マントルの上昇流をホットプルームと呼ぶ垂直対流によって循環
コア(外核にある液体の鉄)の対流が地球磁場の形成の原因とするダイナモ説は、未解明である
地殻と硬いマントルの最上部をプレート、プレートを動かすマントルの水平対流とプレートの
下降流をコールドプルーム、マントルの上昇流をホットプルームと呼ぶ垂直対流によって循環
コア(外核にある液体の鉄)の対流が地球磁場の形成の原因とするダイナモ説は、未解明である
深さ: 0 60km 660km 2900km 5100km 6400km
地殻 上部 下部マントル 外核 内核
厚さ: 60km600km 2240km 2200km 1300km
地表:├─────┼─────┼─────┼─────┼─────┼─────┼─○:地球中心
0 1000km 2000km 3000km 4000km 5000km 6000km
地殻 上部 下部マントル 外核 内核
厚さ: 60km600km 2240km 2200km 1300km
地表:├─────┼─────┼─────┼─────┼─────┼─────┼─○:地球中心
0 1000km 2000km 3000km 4000km 5000km 6000km
|
地殻 6km~60km :海底(厚さ6km)で玄武岩、陸地(厚さ30~60km)で花崗岩 地殻とマントル最上部間:リソスフェア(剛性)、厚さ100kmのプレート マントル 60km~2900km:カンラン石など珪酸塩鉱物、深部は相転移 上部 60km~660km :アセノスフェア(柔らかくて流動性がある) 下部 660km~2900km:メソスフェア(高剛性) 外核 2900km~5100km:厚さ2200km、4400℃~6100℃、液体の鉄やニッケル 内核 5100km~6400km:厚さ1300km、5000℃~6000℃、固体の鉄やニッケル |
|
岩石組成 酸素・ケイ素・アルミ二ウム・マグネシウム・カルシウム・鉄の6元素98% 大気組成 窒素:酸素≒4:1(合計で99%)、その他は少量のアルゴン・二酸化炭素等 海水成分 塩化ナトリウム、塩化マグネシウム、硫酸マグネシウム、硫酸カルシウム等 海流 表層海流:偏西風や貿易風が原動力 深層海流:海水の密度(温度や塩分)の違いが原動力 海洋大循環:表層海流→深層海流→表層海流→深層海流、と循環している |
|
プレート 岩盤、地殻と最上部マントル(リソスフェア)、大陸プレート、海洋プレート プレート同士が遠ざかる場所:海嶺(新たな海洋地殻)、地溝帯、火山活動 プレート同士が近づく場所 :山脈・海溝(深い海底)、地震、 火山活動 プレート同士がすれ違う場所:トランスフォーム断層(横ずれ)、内陸地震 断層 正断層(引っ張り、海嶺)、逆断層(押し合い、海溝)、横ずれ断層(水平移動) 火山活動 マントル上昇マグマ :海嶺沿いの火山、海底火山 沈み込み帯マグマ :海溝沿いの火山 ホットスポットマグマ:ホットプルームマグマによる火山、火山列ができる 地震 プレート境界型地震:沈み込み帯、海溝、固着域ゆがみ復元時に地震が発生 プレート内地震:活断層が動く(ずれる)ことで発生 スラブ内地震 :沈み込み後の海洋プレート(スラブ)内部で発生 地震波 :P波(速い,固体や液体中)、S波(遅い,固体中) 地殻:固体、マントル:固体、外殻:液体、内核:固体 マグニチュード:地震の大きさ、1増えると地震のエネルギーが32倍になる 震度 :揺れの強さ、10階級(0~4,5弱,5強,6弱,6強,7) |
誕生したばかりの地球には微惑星や隕石の衝突のエネルギーで地表は溶けたマグマオーシャン
で覆われ、放出されたガスで水素・メタン・アンモニアからなる原始大気が作られた
火星サイズの原始惑星が斜めに衝突して地球マントルと衝突天体の一部が合体して月ができた
微惑星中の水蒸気や水を含む彗星のような小天体から供給された水蒸気が冷えて海ができた
岩石蒸気と水蒸気でなる原始大気は、鉱物凝縮雨や水蒸気凝縮熱雨による水溶解成分除去後に
水素・二酸化炭素・一酸化炭素・窒素でなる初期大気に変わる(後に光合成生物が酸素を放出)
で覆われ、放出されたガスで水素・メタン・アンモニアからなる原始大気が作られた
火星サイズの原始惑星が斜めに衝突して地球マントルと衝突天体の一部が合体して月ができた
微惑星中の水蒸気や水を含む彗星のような小天体から供給された水蒸気が冷えて海ができた
岩石蒸気と水蒸気でなる原始大気は、鉱物凝縮雨や水蒸気凝縮熱雨による水溶解成分除去後に
水素・二酸化炭素・一酸化炭素・窒素でなる初期大気に変わる(後に光合成生物が酸素を放出)
|
大気圏:下層→対流圏・成層圏・中間圏・熱圏・外気圏←上層、オゾン層は成層圏の中 11km 50km 80km 800km 1万km 電離層は熱圏~中間圏 |
全球凍結(スノーボールアース):地球全体が赤道付近も含め完全に氷床や海氷に覆われた状態、脱出は大規模な火山噴火
大規模な火山噴火→二酸化炭素の大量放出→温暖化加速→激しい雨風→陸地岩石中の栄養塩が海へ→光合成生物の大繁殖
大酸化イベント:大繁殖した光合成生物の酸素放出による大気中の酸素濃度の急激な上昇、豊富な酸素がオゾン層を生成
オゾン層は、太陽からの有害な波長の紫外線の多くを吸収し、地上の生態系を保護する役割を果たしている
┌→停滞→二酸化炭素の供給減少→寒冷化→化学反応速度低下→全球凍結
火山活動─┤
└→活発→二酸化炭素の供給増加→温暖化→化学反応速度増大→大酸化イベント
┌─────────炭素循環←──────────┐
↓ │
┌─二酸化炭素の消費─┐ ┌─化学風化作用─┐ 二酸化炭素の供給
↓ ↓ ↓ ↓ ↑
「光合成→有機物の堆積、雨水→炭酸+地表岩石→炭酸塩鉱物」→大規模な火山噴火
│↑ │ ↑
│└──────光合成生物の大繁殖←─────┘ 全球凍結
↓ ↑
酸素濃度の上昇 火山活動停滞・二酸化炭素の供給減少→寒冷化の暴走
↓
縞状鉄鉱層の形成(海中)→赤色砂岩を形成(地表)→大気中へ放出(成層圏)→オゾン層を生成
|
40億年前 27億年前 21億年前 12億年前 生命の誕生───→藍藻類の出現───→真核生物の誕生───→真核生物のビッグバン 真正細菌 酸素をつくる光合成 ミトコンドリアの出現 葉緑体の出現 |
|
※酸素を放出する光合成生物であるシアノバクテリアは、原核生物の一種、真正細菌である ※真核生物は古細菌に真正細菌が細胞内共生して誕生したと考えられている(古細菌が宿主) ※20億年前プロテオバクテリアが出現し真核細胞内に共生してミトコンドリアへと進化した ※真核生物のビッグバン:爆発的に進化・多様化、多数の分類群に急激に分化 |
| 嫌気性原核生物(A) |
酸素呼吸する ミトコンドリア 好気性原核生物(B) |
べん毛を持つ 原核生物(C) |
光合成する シアノバクテリア 原核生物(D) | |
| 原核生物 | (A) | |||
| 真核生物 | (A+B) | |||
| (A+B+C) | ||||
| ミドリムシ | (A+B+C+D) |
|
シアノバクテリアの祖先は30~25億年前に地球上に出現し、初めて酸素発生型光合成を始めました 光合成では水を電子供与体とすることができるため、 水と光があればエネルギーが得られることとなり、当時の地球上で大繁殖したようです (水分子から電子を奪い、その副産物として酸素ができます) ←↑:大酸化イベント |
|
葉緑体の起源はシアノバクテリア、ミトコンドリアの起源はプロテオバクテリア(好気性細菌) という原核生物で、それぞれ独自のDNAを持ち、分裂して増えていた生き物です |
|
植物細胞の中には、光合成によって、光エネルギーから有機物を合成する葉緑体と、 呼吸によって、有機物が蓄えた化学エネルギーをATPとして取り出すミトコンドリアが存在します |
|
光合成の電子伝達系により、光エネルギーをATPやNADPHといった化学エネルギーに変換します 次いで、葉緑体で、その化学エネルギーをもとに二酸化炭素を固定し、有機化合物を合成します |
|
ミトコンドリアで行われている呼吸では、酸素を用いて、有機物を二酸化炭素と水に分解します その過程で、生命活動に必要なエネルギーをATPとして取り出しています |
|
植物の還元反応:太陽光線→水から水素を奪う+二酸化炭素→酸素+炭水化物 葉緑体の機能 動物の酸化反応:炭水化物+水素を奪う酸素を吸う→二酸化炭素+水 ミトコンドリアの機能 |
|
還元:左→右 6H2O+6CO2⇔6O2+C6H12O6 酸化:左←右 |
|---|
|
・細胞内共生説とは、真核生物が原核生物と共生することによって、 細胞小器官を持った細胞を形成したという仮説です ・植物細胞は、ミトコンドリアと葉緑体という細胞小器官を持っています 実は、この2器官はそれぞれ独立して生きていたバクテリアで、 葉緑体は光合成するシアノバクテリア、ミトコンドリアは呼吸をする プロテオバクテリアとして存在していました |
|
・約20億年前に、ある真核生物がプロテオバクテリアを取り込みました 真核生物の細胞の中で、プロテオバクテリアがミトコンドリアへと変化し、 菌類や動物へと進化しました |
|
・一方、約10億年前にプロテオバクテリアを取り込んだ細胞が、シアノ バクテリアを取り込み、シアノバクテリアが葉緑体へと変化して、 藻類や植物へと進化を遂げました |
|
・共生する相手を取り込んだ生物を宿主(しゅくしゅ)と呼びます 共生生物は宿主からは好適な生育環境を得ていると考えられている ・細胞内共生とは、宿主の生物が異なる種の原核生物を取り込んで共生し、 細胞小器官を持った細胞を形成したことなのです 真核生物は古細菌に真正細菌が細胞内共生して誕生したと考えられている 宿主の増殖に合わせて共生細胞の増殖は宿主にコントロールされています 宿主と葉緑体やミトコンドリアは切り離せない関係(絶対共生)になった |
|
・葉緑体やミトコンドリアがエネルギー変換の際に活性酸素を放出するので、 原核生物が宿主と共生して細胞小器官へと進化する以前に、宿主にとって 猛毒の活性酸素から遺伝子を守るために核膜をもつようになった |
|
40億年前 23億年前 21億年前 7億年前 6億年前 原核生物─→全球凍結─→真核生物─→全球凍結─→多細胞生物 1~10μm 5~100μm 2億年前:シアワセモが現存最小 単細胞のみ 単細胞:1mm以下 多細胞:20μm~30μm |
単細胞生物は、酸素を利用してコラーゲンを生成し、細胞同士を結合させて多細胞生物に進化
単細胞生物が進化して定数群体をなし、細胞が分化して、内外組織や器官を形成するに至った
単細胞生物(植物性・中間的・動物性)→多細胞生物(植物・菌類・動物)→エディアカラ生物群
単細胞生物が進化して定数群体をなし、細胞が分化して、内外組織や器官を形成するに至った
単細胞生物(植物性・中間的・動物性)→多細胞生物(植物・菌類・動物)→エディアカラ生物群
ディッキンソニア、ネミアナ、チャルニア、キンベレラ、スプリッギナ、シクロメデューサ
トゥリプラキディウム、フィロゾーン、スワートブンティア、エルニエッタ、ヨルギア
トゥリプラキディウム、フィロゾーン、スワートブンティア、エルニエッタ、ヨルギア
カンブリア大爆発とは、一気に一万種類の水生動物が出現した化石記録(パージェス頁岩動物
群化石やチェンジャン動物群化石)の爆発的多様化であり、進化的な爆発は3億年前からで、
要因は捕食関係が生まれ、淘汰圧による生存競争の勝利と言う
遺伝子の爆発的多様化(進化的な爆発) 化石記録の爆発的多様化
─┼───────┼──────┼───────┼─────┼────
3億年前 カンブリア紀第3期
カンブリア紀の最初の覇者はアノマロカリスや三葉虫など硬い殻と複眼を持つ節足動物である
カンブリア紀:アノマロカリス、アイシュアイア、ウィクシア、ピカイア、マルレラ
チャンセロリア、ワプティア、レアンコイリア、フクシアンフィア
エルドニア、イソキシス、オパビニア、ハルキゲニア、三葉虫
群化石やチェンジャン動物群化石)の爆発的多様化であり、進化的な爆発は3億年前からで、
要因は捕食関係が生まれ、淘汰圧による生存競争の勝利と言う
遺伝子の爆発的多様化(進化的な爆発) 化石記録の爆発的多様化
─┼───────┼──────┼───────┼─────┼────
3億年前 カンブリア紀第3期
カンブリア紀の最初の覇者はアノマロカリスや三葉虫など硬い殻と複眼を持つ節足動物である
カンブリア紀:アノマロカリス、アイシュアイア、ウィクシア、ピカイア、マルレラ
チャンセロリア、ワプティア、レアンコイリア、フクシアンフィア
エルドニア、イソキシス、オパビニア、ハルキゲニア、三葉虫
オルドビス紀:三葉虫、オウムガイ、ウミサソリ、筆石、ウミユリ、床板サンゴ
シルル紀 :ウミサソリ
デボン紀 :トビムシ、ウミサソリ、アンモナイト(ペルム紀に絶滅後ジュラ紀に再繁栄)
石炭紀 :メガネウラ、アースロプレウラ
シルル紀 :ウミサソリ
デボン紀 :トビムシ、ウミサソリ、アンモナイト(ペルム紀に絶滅後ジュラ紀に再繁栄)
石炭紀 :メガネウラ、アースロプレウラ
カンブリア紀に脊椎を獲得したピカイア・コノドント→顎口類→硬骨魚類→肉鰭類→両生類
絶滅魚類:板皮類(ダンクルオステウス)、棘魚類(顎をもつ最初の脊椎動物、ヒレは硬い棘)
絶滅魚類:板皮類(ダンクルオステウス)、棘魚類(顎をもつ最初の脊椎動物、ヒレは硬い棘)
┌無顎類(円口類)─ヤツメウナギ、ヌタウナギ──┐
│ │
脊椎動物┤ ┌軟骨魚類─サメ、エイ │
│ │ │
└顎口類┤ ┌条鰭類─コイ、タイ │魚類:四足類を除く左記全て
│ │ │
└硬骨魚類┤ ┌シーラカンス類 │
│ │ │
└肉鰭類┼肺魚類─ハイギョ──┘
│
└四足類┬両生類
│
└羊膜類─爬虫類、鳥類、ほ乳類
│ │
脊椎動物┤ ┌軟骨魚類─サメ、エイ │
│ │ │
└顎口類┤ ┌条鰭類─コイ、タイ │魚類:四足類を除く左記全て
│ │ │
└硬骨魚類┤ ┌シーラカンス類 │
│ │ │
└肉鰭類┼肺魚類─ハイギョ──┘
│
└四足類┬両生類
│
└羊膜類─爬虫類、鳥類、ほ乳類
| 生物 | 陸上進出 |
|---|---|
| 植物の陸上進出 | クックソニアは胞子嚢により空気中でも繁殖可能とし、乾燥対策にクチクラ層という膜で表面を覆う |
| コケ類などの陸上進出後は、植物は根・茎・葉の通道組織の役目の維管束を発達させ、巨木となった | |
| 昆虫の陸上進出 | 体の小さい昆虫は世代交代が短期間なので進化が速く、加えて泳ぐためのエラを空を飛ぶ翅に変えた |
| 昆虫は少ない食物を有効活用でき、食べ物を変えることで同じ環境で複数の種が暮らせる利点がある | |
| 動物の陸上進出 | 両生類が、肺・腎臓・骨と筋肉をもつ足・卵を乾燥から守る羊膜を獲得して水中から陸上に進出した |
| 鳥類の祖の恐竜は低酸素でも効率的な呼吸ができる気嚢を獲得して肺呼吸と違う呼吸システムに進化 |
植物体全体の姿はわかりません
陸上植物の全体の姿が残された最も古い化石が、古生代シルル紀中期(約4億2500万年前)のクックソニアです
これはリニア植物の一種で、根も葉も無く、先端に胞子の入った袋を持ち、水の通る管を持っていました
リニア植物は、古生代デボン紀にかけて繁茂しましたが、絶滅しました
地球の歴史上最初の陸生動物は節足動物門のウミサソリ類だったと考えられている(両生類と同時期かそれよりも早く)
PT境界大量絶滅を生き延びた単弓類の一類が哺乳形類に進化した
哺乳類の祖は、以下の真の哺乳類の特徴のいくつかを備えていた
内温性、鼻孔と口腔を隔てる骨、横隔膜、下顎の骨、耳小骨3つ
哺乳類の祖は、以下の真の哺乳類の特徴のいくつかを備えていた
内温性、鼻孔と口腔を隔てる骨、横隔膜、下顎の骨、耳小骨3つ
三畳紀は、単弓類、クルロタルス類(双弓類)、恐竜が生存競争を繰り広げる
PT境界大量絶滅を生き残った単弓類リストロサウルスも後に恐竜に代られる
TJ境界大量絶滅では多くのクルロタルス類は絶滅するが、ワニ形類は生き延
びてジュラ紀に台頭した恐竜と生存競争、単弓類のキノドン類は小型化
クルロタルス類の代表格はサウロスクス、ワニ形類の代表格はプロトスクス
PT境界大量絶滅を生き残った単弓類リストロサウルスも後に恐竜に代られる
TJ境界大量絶滅では多くのクルロタルス類は絶滅するが、ワニ形類は生き延
びてジュラ紀に台頭した恐竜と生存競争、単弓類のキノドン類は小型化
クルロタルス類の代表格はサウロスクス、ワニ形類の代表格はプロトスクス
硬いヒレをもつ肉鰭類→両生類→羊膜類(羊膜と卵殻をもつ四肢動物)→単弓類(爬虫類とは別物)→獣弓類→哺乳形類
両生類は古生代石炭紀に哺乳類の祖先である単弓類と恐竜の祖先である爬虫類に分かれ、恐竜は一部が残って鳥類に進化
TJ境界大量絶滅を生き延びた恐竜形類:小型で直立歩行する爬虫類のトカゲで、俊敏性と内温性を備える優れた体の構造
両生類は古生代石炭紀に哺乳類の祖先である単弓類と恐竜の祖先である爬虫類に分かれ、恐竜は一部が残って鳥類に進化
TJ境界大量絶滅を生き延びた恐竜形類:小型で直立歩行する爬虫類のトカゲで、俊敏性と内温性を備える優れた体の構造
両生類──羊膜類─┬─単弓類─┬─盤竜類:ディメトロドン
イクチオステガ │ │
│ └─獣弓類:エステメノスクス,キノドン類:哺乳型類(哺乳類の祖先)
│
│ ┌─無弓類:側頭窓がない(単弓類の側頭窓は哺乳類の耳の穴の起源)
│ │
└─竜弓類───爬虫類─┴─双弓類:恐竜・ワニ・トカゲの祖先
恐竜:三畳紀に出現、白亜紀に鳥類以外絶滅
イクチオステガ │ │
│ └─獣弓類:エステメノスクス,キノドン類:哺乳型類(哺乳類の祖先)
│
│ ┌─無弓類:側頭窓がない(単弓類の側頭窓は哺乳類の耳の穴の起源)
│ │
└─竜弓類───爬虫類─┴─双弓類:恐竜・ワニ・トカゲの祖先
恐竜:三畳紀に出現、白亜紀に鳥類以外絶滅
┌─獣脚類 ティラノサウルス 肉食 羽毛を持つものが多い
│ ヴェロキラプトル 肉食
│ デイノニクス 肉食
│
┌─竜盤類─┴─竜脚形類 アンペロサウルス 植物食
│ ブラキオサウルス 植物食
│ ブラテオサウルス 植物食
│ ディプロドクス 植物食
│
┌─恐竜類─┤ ┌─装盾類 アンキロサウルス 植物食 鎧竜類
│ │ │ エウオプロケファルス植物食 鎧竜類
│ │ │ ケントロサウルス 植物食 剣竜類
│ │ │
│ └─鳥盤類─┼─周飾頭類 トリケラトプス 植物食 角竜類
双弓類──主竜類─┤ │ ハキケフォラサウルス植物食 堅頭竜類
恐竜様類│ │
恐竜形類│ └─鳥脚類 パラサウロロフス 植物食
│ コリトサウルス 植物食
│
└─────────────翼竜類 プテラノドン 魚食
ランフォリンクス 魚食
その他───────────────首長竜 プレシオサウルス 魚食
魚竜 ステノプテリギウス 肉食
恐竜絶滅直後の生き残り 恐鳥類 ガストルニス 肉食
※恐竜は現在の鳥類の祖先にあたる獣脚類の一部の系統を残して、白亜紀末に絶滅した
│ ヴェロキラプトル 肉食
│ デイノニクス 肉食
│
┌─竜盤類─┴─竜脚形類 アンペロサウルス 植物食
│ ブラキオサウルス 植物食
│ ブラテオサウルス 植物食
│ ディプロドクス 植物食
│
┌─恐竜類─┤ ┌─装盾類 アンキロサウルス 植物食 鎧竜類
│ │ │ エウオプロケファルス植物食 鎧竜類
│ │ │ ケントロサウルス 植物食 剣竜類
│ │ │
│ └─鳥盤類─┼─周飾頭類 トリケラトプス 植物食 角竜類
双弓類──主竜類─┤ │ ハキケフォラサウルス植物食 堅頭竜類
恐竜様類│ │
恐竜形類│ └─鳥脚類 パラサウロロフス 植物食
│ コリトサウルス 植物食
│
└─────────────翼竜類 プテラノドン 魚食
ランフォリンクス 魚食
その他───────────────首長竜 プレシオサウルス 魚食
魚竜 ステノプテリギウス 肉食
恐竜絶滅直後の生き残り 恐鳥類 ガストルニス 肉食
※恐竜は現在の鳥類の祖先にあたる獣脚類の一部の系統を残して、白亜紀末に絶滅した
| 地層 | 年代 | 出来事 | ||
|---|---|---|---|---|
| 表土 | 現在 | |||
| 1 | カイバブ石灰岩層 | 2億6千万年前 | 恐竜はこの地層で繁栄と絶滅 | |
| 2 | トロウィープ層 | 2億6千2百万年前 | ||
| 3 | ココニノ砂岩 | 2億6千5百万年前 | グランドキャニオンが陸地になる | |
| 4 | ハーミット泥板岩 | 2億7千万年前 | ||
| 5 | スーパイ・グループ | 2億7千5百万年前 | ||
| 6 | レッドウォール石灰岩 | 3億4千万年前 | 爬虫類の出現 | |
| 7 | テンプル・ビュート石灰岩 | 3億7千万年前 | 脊椎動物の上陸・両生類の出現 | |
| 8 | ムアヴ石灰岩 | 5億年前 | 植物の上陸 | |
| 9 | ブライトエンジェル泥板岩 | 5億1千万年前 | ||
| 10 | テーピーツ砂岩 | 5億2千万年前 | カンブリア生命大爆発 | |
| 11 | グランドキャニオンスーパーグループ(4層) | 7億4千万年前 | 全球凍結 | |
| 12 | ヴィシュヌ基盤岩(花崗岩変成岩帯) | 16億8千万年前 | 激しい火山活動と巨大大陸の形成 | |
| コロラド川 | コロラド川が浸食中 | |||
恐竜絶滅後の地球上で哺乳類の進化が始まりました(授乳は哺乳類の特徴の一つで名前の由来です)
新生代の期間は中生代の白亜紀より短いのですが、哺乳類はその短期間のうちに進化・多様化します
このため、生存競争も激しく、進化と同時に絶滅種も出てきます
デスモスチルスは新生代中新世に生息していた大型の哺乳類ですが、その後絶滅してしまいます
新生代の期間は中生代の白亜紀より短いのですが、哺乳類はその短期間のうちに進化・多様化します
このため、生存競争も激しく、進化と同時に絶滅種も出てきます
デスモスチルスは新生代中新世に生息していた大型の哺乳類ですが、その後絶滅してしまいます
┌─単孔類:カモノハシ、ハリモグラ
│
哺乳類─┤ ┌─有袋類:オポッサム、カンガルー
│ │
└─┤ ┌─アフリカ獣類:アフリカ食虫類、近蹄類
│ │
└─有胎盤類─┼─異節類:ナマケモノ、アリクイ、アルマジロ
│
└─北方真獣類:サル(霊長類)、ネズミ、ウサギ、コウモリ、ウマ、
サイ、バク、ラクダ、ウシ、クジラ、ネコ
│
哺乳類─┤ ┌─有袋類:オポッサム、カンガルー
│ │
└─┤ ┌─アフリカ獣類:アフリカ食虫類、近蹄類
│ │
└─有胎盤類─┼─異節類:ナマケモノ、アリクイ、アルマジロ
│
└─北方真獣類:サル(霊長類)、ネズミ、ウサギ、コウモリ、ウマ、
サイ、バク、ラクダ、ウシ、クジラ、ネコ
猿類→類人猿→人類
霊長類─┬→オモミス類─→直鼻猿類─┬→真猿類──┬→狭鼻猿類⇒
│ │ │
└→アダピス類─→曲鼻猿類 └→メガネザル└→広鼻猿類
┼─────┼──────┼──────┼──────┼────→
7500 6550 6300 5800 4000 万年前
⇒┬→ヒト上科─┬→ヒト科──┬→ヒト亜科──┬→ヒト族─┬→ヒト亜族─→ヒト属
│ │ │ │ │
└→オナガザル└→テナガザル└→オランウータン└→ゴリラ └→チンパンジー
───┼──────┼──────┼───────┼─────┼─────┼────
2500 1800 1200 700 600 240万年前
広鼻猿類:新世界ザル、オナガザル:旧世界ザル、ヒト上科/ヒト科:類人猿、ヒト亜族/ヒト属:人類│ │ │
└→アダピス類─→曲鼻猿類 └→メガネザル└→広鼻猿類
┼─────┼──────┼──────┼──────┼────→
7500 6550 6300 5800 4000 万年前
⇒┬→ヒト上科─┬→ヒト科──┬→ヒト亜科──┬→ヒト族─┬→ヒト亜族─→ヒト属
│ │ │ │ │
└→オナガザル└→テナガザル└→オランウータン└→ゴリラ └→チンパンジー
───┼──────┼──────┼───────┼─────┼─────┼────
2500 1800 1200 700 600 240万年前
二足歩行:恐竜類、鳥類(翼に変わった前肢→二足歩行)、霊長類以外の哺乳類ではカンガルーのみ
霊長類の二足歩行:樹上生活の進化→地上生活→類人猿:普段は四足歩行(ナックル・ウォーキング)
ヒト科:類人猿、初期猿人→猿人→原人→旧人→新人(現生人類)、安定した直立二足歩行は猿人から
二足歩行の最大の欠点は走るのが遅いこと、直立二足歩行にはメリットがあるが、デメリットもある
人類の出現:700万年前、サヘラントロプス=チャデンシス「トゥーマイ=生命の希望」猿人
霊長類の二足歩行:樹上生活の進化→地上生活→類人猿:普段は四足歩行(ナックル・ウォーキング)
ヒト科:類人猿、初期猿人→猿人→原人→旧人→新人(現生人類)、安定した直立二足歩行は猿人から
二足歩行の最大の欠点は走るのが遅いこと、直立二足歩行にはメリットがあるが、デメリットもある
人類の出現:700万年前、サヘラントロプス=チャデンシス「トゥーマイ=生命の希望」猿人
アルディピテクス・ラミダス ラミダス猿人 直立二足歩行
↓
アウストラロピテクス・アファレンシス アファール猿人 ルーシー、現生人類の祖先
↓
ホモ・ハピリス 能力ある人(原人) 旧石器時代
↓
ホモ・エレクトゥス 直立する人(原人) ジャワ原人・北京原人
↓
ホモ・ハイデルベルゲンシス ハイデルベルクの人(旧人) ネアンデルタール人と現生人類の祖先
↓
ホモ・サピエンス 賢い人(新人) クロマニヨン人、現生人類
上記の他に、オロリン属1種、アルディピテクス属1種、ケニアントロプス属1種、
アウストラロピテクス属4種、パラントロプス属3種、ホモ属3種、などがある
↓
アウストラロピテクス・アファレンシス アファール猿人 ルーシー、現生人類の祖先
↓
ホモ・ハピリス 能力ある人(原人) 旧石器時代
↓
ホモ・エレクトゥス 直立する人(原人) ジャワ原人・北京原人
↓
ホモ・ハイデルベルゲンシス ハイデルベルクの人(旧人) ネアンデルタール人と現生人類の祖先
↓
ホモ・サピエンス 賢い人(新人) クロマニヨン人、現生人類
上記の他に、オロリン属1種、アルディピテクス属1種、ケニアントロプス属1種、
アウストラロピテクス属4種、パラントロプス属3種、ホモ属3種、などがある
| 地質時代 | 考古年代 | 道具・生活・住居・習俗 | 人類・社会・文明 | |||
|---|---|---|---|---|---|---|
| 中新世 鮮新世 | 先史時代 | 700万年前 | 直立二足歩行 初期の打製石器、礫石器の使用 狩猟・採集・漁労(獲得経済)、群社会 | 猿人 | トゥーマイ猿人 ラミダス猿人 アファール猿人 | |
| 更新世 | 260万年前 ~30万年前 | 前期 旧石器時代 | 火の使用 石核石器の発達(握斧・尖頭器) 言語の使用、洞穴住居 | 原人 | ホモ・ハピリス ホモ・エレクトゥス ジャワ原人、北京原人 | |
| 30万年前 ~3万年前 | 中期 旧石器時代 | 剥片石器の発達 死者の埋葬始まる、衣服の着用(獣皮) | 旧人 | ハイデルベルク人 ネアンデルタール人 | ||
| 3万年前 ~1万年前 | 後期 旧石器時代 | 骨角器の製作(槍・銛・釣針・針) ランプの使用 装身具の使用(石・貝・牙・角) 洞穴絵画(アルタミラ・ラスコー) 女性裸像、呪術的行為の発生 屈葬・副葬品 | 新人 | クロマニヨン人 グリマルディ人 周口店上洞人、浜北人 ホモ・サピエンス | ||
| 完新世 | 1万年前 | 中石器時代 | 細石器の製作(鏃・銛・釣針) 漁労技術の進歩(釣針・網) 弓矢の使用、そり・車の発明、岩絵 | 動植物の家畜化 | ||
| 9000年前 | 新石器時代 | 農耕・牧畜(生産経済) 磨製石器の製作(石斧・石鎌・石包丁) 土器の普及(彩文土器) 織物の始まり(羊毛・麻) 地母神像、竪穴式住居(定住) | 祭祀を中心に集落を形成 氏族・部族社会 | |||
| 原史時代 | 4000年前 | 青銅器時代 | 文字が発明され文字記録が始まる 都市国家の成立 | エジプト文明(象形文字) メソポタミア文明(楔形文字) インダス文明(インダス文字) 黄河・長江文明(甲骨文字) | ||
| 歴史時代 | 3000年前 | 鉄器時代 | 青銅器の製作 巨石建造物(ストーンヘンジなど) | |||
25億年前:ケノリア超大陸(地層が少なく検証が難しい)
19億年前:ヌーナ超大陸→誕生直後から分裂始める
18億年前:コロンビア超大陸→15億年前に分裂
15億年前:パノティア超大陸→10億年前に分裂
10億年前:ロディニア超大陸→6億年前にゴンドワナ大陸・ローレンシア大陸などに分裂
3億年前:パンゲア超大陸の誕生(パンゲア:全ての陸地、パンサラッサ:全ての海という意味)
約1億7500万年前、超大陸パンゲアは北がローラシア大陸に南がゴンドワナ大陸に分裂、ローラシア
大陸は西が北米大陸に東がユーラシア大陸に分裂して北大西洋が誕生、1億2000万年前、ゴンドワナ
大陸から、南極大陸とオーストラリア大陸が分かれて南に移動、ゴンドワナ大陸は、南米大陸の東海
岸とアフリカ大陸の西海岸が裂けて南大西洋になり、9000万年前、インド大陸は、アフリカ大陸から
分かれて北上、5500万年前にユーラシア大陸と衝突したため、ユーラシア大陸側が大規模に隆起して
ヒマラヤ山脈が誕生した
始新世:南アメリカから南極とオーストラリアが分裂
漸新世直前:ロッキー山脈、アンデス山脈が形成された
アフリカ大陸もユーラシア大陸西側の欧州大陸と衝突して、アルプス山脈が形成された
19億年前:ヌーナ超大陸→誕生直後から分裂始める
18億年前:コロンビア超大陸→15億年前に分裂
15億年前:パノティア超大陸→10億年前に分裂
10億年前:ロディニア超大陸→6億年前にゴンドワナ大陸・ローレンシア大陸などに分裂
3億年前:パンゲア超大陸の誕生(パンゲア:全ての陸地、パンサラッサ:全ての海という意味)
約1億7500万年前、超大陸パンゲアは北がローラシア大陸に南がゴンドワナ大陸に分裂、ローラシア
大陸は西が北米大陸に東がユーラシア大陸に分裂して北大西洋が誕生、1億2000万年前、ゴンドワナ
大陸から、南極大陸とオーストラリア大陸が分かれて南に移動、ゴンドワナ大陸は、南米大陸の東海
岸とアフリカ大陸の西海岸が裂けて南大西洋になり、9000万年前、インド大陸は、アフリカ大陸から
分かれて北上、5500万年前にユーラシア大陸と衝突したため、ユーラシア大陸側が大規模に隆起して
ヒマラヤ山脈が誕生した
┌─西→北米大陸
┌─北→ローラシア大陸──┤
| └─東→ユーラシア大陸←───衝突←──┐
| ↑
超大陸パンゲア──┤ ┌─西→南米大陸 北上
の分裂 | ┌─北──┤ ↑
| | └─東→アフリカ大陸、インド大陸
└─南→ゴンドワナ大陸──┤
|
└─南→南極大陸、オーストラリア大陸
暁新世:アフリカと南アメリカが分離、アフリカからインドが離れた┌─北→ローラシア大陸──┤
| └─東→ユーラシア大陸←───衝突←──┐
| ↑
超大陸パンゲア──┤ ┌─西→南米大陸 北上
の分裂 | ┌─北──┤ ↑
| | └─東→アフリカ大陸、インド大陸
└─南→ゴンドワナ大陸──┤
|
└─南→南極大陸、オーストラリア大陸
始新世:南アメリカから南極とオーストラリアが分裂
漸新世直前:ロッキー山脈、アンデス山脈が形成された
アフリカ大陸もユーラシア大陸西側の欧州大陸と衝突して、アルプス山脈が形成された
新生代初期の温暖化:暁新世から始新世にかけて1~2万年の短期間
新生代の寒冷化 :始新世/漸新世境界と中新世後期の寒冷化
第四紀は10万年周期の氷期・間氷期繰り返し
最終氷期は1500年周期の急激な温暖化が繰り返される
新生代の寒冷化 :始新世/漸新世境界と中新世後期の寒冷化
第四紀は10万年周期の氷期・間氷期繰り返し
最終氷期は1500年周期の急激な温暖化が繰り返される
| 予想変動 | 予想被害 |
|---|---|
| 太陽フレアの増加による磁気嵐 | 通信障害や広域停電 |
| 地磁気の減少による宇宙線の増加 | 雲による(日傘効果)で寒冷化 |
| 隕石衝突被害 | 衝撃波による人的・物的被害 |
| 巨大地震 | 震災、火災、津波 |
| 火山の巨大噴火 | 溶岩流、火山の冬 |
| 温暖化による自然災害の増加 | 熱波、干ばつ、暴風雨、高潮、洪水 |
| 環境変動 | 社会的リスク |
|---|---|
| 湿潤地域の降水量の増加 | 食料や水資源の確保の問題、 人々の強制移転の増加、 経済成長の減速、 貧困の増加、 内戦や民族紛争 |
| 乾燥地域の乾燥化 | |
| 海面水位上昇、海洋酸性化 | |
| 海氷・氷床の減少、永久凍土の減少 | |
| 極端な気象現象の頻度の増加 | |
| 生物種の絶滅、生態系の遷移 | |
| 世界平均気温の上昇幅 | |
| 厳しい二酸化炭素排出緩和 → 0.3~1.7度 | |
| 排出抑制の追加的努力なし → 2.6~4.8度 | |
| 地質時代境界 | 年代 | 事件 | 原因 | 代表的な絶滅種 |
|---|---|---|---|---|
| オルドビス紀|シルル紀 | 4億4340万±150万年前 | OS境界大量絶滅 | ||
| デボン紀|石炭紀 | 3億5890万±40万年前 | FF境界大量絶滅 | 海洋無酸素イベント | |
| ペルム紀|三畳紀 | 2億5217万±6万年前 | PT境界大量絶滅 | 海洋無酸素イベント | 三葉虫 |
| 三畳紀|ジュラ紀 | 2億130万±2万年前 | TJ境界大量絶滅 | 火山活動活発化 | |
| 白亜紀|古第三紀 | 6550万年前 | KPg境界大量絶滅 | 小惑星の衝突 | アンモナイト,恐竜 |
絶滅の原因?:コールドプルームの落下→スーパープルームの上昇→大規模火成活動→温暖化と海洋無酸素イベント
海洋無酸素イベント:沈殿有機物が海水溶存酸素と反応して二酸化炭素を放出、超温暖化により陸上及び海洋生物が死滅
上記5回の大量絶滅の前後に2回と、上記前部4回の各中間に3回の、計5回若干規模の小さい絶滅があったとされるが
小天体の重爆撃・大酸化イベント・全球凍結・超大陸の形成と分裂時のスーパープルームでも大量絶滅があったとされる
海洋無酸素イベント:沈殿有機物が海水溶存酸素と反応して二酸化炭素を放出、超温暖化により陸上及び海洋生物が死滅
上記5回の大量絶滅の前後に2回と、上記前部4回の各中間に3回の、計5回若干規模の小さい絶滅があったとされるが
小天体の重爆撃・大酸化イベント・全球凍結・超大陸の形成と分裂時のスーパープルームでも大量絶滅があったとされる
| 地質時代 | 開始年代 | 出来事 | 動物界 | 植物界 | |
|---|---|---|---|---|---|
| 先カンブリア時代 | 冥王代 | 45.67億年前 | 地球の誕生、小天体の重爆撃、月の誕生 原始大気と原始海洋の出現 | 有機化合物の合成 始原生物(共通祖先)の出現 | |
| 太古代 | 40.31億年前 | 陸地の出現、小天体後期重爆撃 地磁気の形成、光合成生物の進化 | 原核生物:真正細菌、古細菌 シアノバクテリア(酸素発生型光合成) | ||
| 原生代 | 25億年前 | 全球凍結、大酸化イベント 縞状鉄鉱層の形成、エディアカラ生物群 | 真核生物、グリパニア 多細胞生物の出現 | ||
| 顕生代 | 古生代 | 5億3880万年前 | カンブリア大爆発、生物大放散事変 生物(植物→昆虫→動物)の陸上進出 | 三葉虫の出現 魚類、両生類 | シダ類森林 裸子植物の出現 |
| 中生代 | 2億5190万2000年前 | アンモナイトの繁栄、始祖鳥 恐竜の繁栄、小惑星の衝突、恐竜の絶滅 | 爬虫類の繁栄 哺乳類の祖が出現 | 被子植物の出現 (虫媒花の進化) | |
| 新生代 | 6600万年前 | 霊長類の進化(猿類→類人猿) 人類の出現 | 飛ぶ鳥の繁栄 哺乳類の繁栄 | 被子植物の繁栄 | |
| 地質時代 | 開始年代 | 出来事 | ||
|---|---|---|---|---|
| 新生代 | 古第三紀 | 暁新世 | 6600万年前 | 大型恐鳥類 |
| 始新世 | 5600万年前 | 小型哺乳類 | ||
| 漸新世 | 3390万年前 | 哺乳類の多様化 | ||
| 新第三紀 | 中新世 | 2303万年前 | アルプス山脈形成 | |
| 鮮新世 | 533万3000年前 | 原始人類 | ||
| 第四紀 | 更新世 | 258万年前 | 石器時代 | |
| 完新世 | 1万1700年前 | 農耕牧畜時代 | ||
| 地質時代 | 考古年代 | 石器/文明 | ||
|---|---|---|---|---|
| 中新世 鮮新世 | 先史時代 | 700万年前 | 打製石器 礫石器 | |
| 更新世 | 260万年前 ~30万年前 | 前期旧石器時代 | 石核石器 | |
| 30万年前 ~3万年前 | 中期旧石器時代 | 剥片石器 | ||
| 3万年前 ~1万年前 | 後期旧石器時代 | 骨角器 | ||
| 完新世 | 1万年前 | 中石器時代 | 細石器 | |
| 9000年前 | 新石器時代 | 磨製石器 | ||
| 原史時代 | 4000年前 | 青銅器時代 | 文字の発明 四大文明 | |
| 歴史時代 | 3000年前 | 鉄器時代 | ||
TOP|
地球史年表(相似的)
|TOP
←────────────────────宇宙の歴史────────────────────→
←─────地質時代─────→
宇暗星 原←─先カンブリア時代──→顕生代
宙黒の 始冥太 原 古中新
創時誕 太王古 生 生生生
成代生 銀河の形成 宇宙の加速膨張 陽代代 代 代代代
┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─
00 06 12 18 24 30 36 42 48 54 60 66 72 78 84 90 96 102 108 114 120 126 132 138 億年後
138 132 126 120 114 108 102 96 90 84 78 72 66 60 54 48 42 36 30 24 18 12 6 0 億年前
←─────地質時代─────→
宇暗星 原←─先カンブリア時代──→顕生代
宙黒の 始冥太 原 古中新
創時誕 太王古 生 生生生
成代生 銀河の形成 宇宙の加速膨張 陽代代 代 代代代
┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─
00 06 12 18 24 30 36 42 48 54 60 66 72 78 84 90 96 102 108 114 120 126 132 138 億年後
138 132 126 120 114 108 102 96 90 84 78 72 66 60 54 48 42 36 30 24 18 12 6 0 億年前
←────────────────先カンブリア時代───────────────→←顕生代→
45.67億年前 40.31億年前 25億年前 5.4 2.50.7億年前
┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─-
46 44 42 40 38 36 34 32 30 28 26 24 22 20 18 16 14 12 10 8 6 4 2 0 億年前
45.67億年前 40.31億年前 25億年前 5.4 2.50.7億年前
冥王代 太古代 原生代 古 中 新
億年 5.36 15 19.59 2.8691.90.7億年┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─┼─-
46 44 42 40 38 36 34 32 30 28 26 24 22 20 18 16 14 12 10 8 6 4 2 0 億年前
←──────────────────-太古代──────────────────→
40.31億年前 36億年前 32億年前 28億年前
┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼
40.5 39.0 37.5 36 34.5 33.0 31.5 30.0 28.5 27.0 25.5 24.0億年前
40.31億年前 36億年前 32億年前 28億年前
原太古代 | 古太古代 | 中太古代 | 新太古代
億年 4 4 4 3 億年┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼
40.5 39.0 37.5 36 34.5 33.0 31.5 30.0 28.5 27.0 25.5 24.0億年前
←────────────────────────原生代────────────────────────→
25億年前 16億年前 10億年前
┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼
25.0 23.5 22.0 20.5 19.0 17.5 16.0 14.5 13.0 11.5 10.0 8.5 7.0 5.5億年前
25億年前 16億年前 10億年前
古原生代 | 中原生代 | 新原生代
億年 9 6 4.59 億年┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼
25.0 23.5 22.0 20.5 19.0 17.5 16.0 14.5 13.0 11.5 10.0 8.5 7.0 5.5億年前
←────────────────────-古生代-────────────────────→
5.3880億年前 4.8540億年前 4.43億年前4.1920億年前 3.5890億年前 2.9890億年前
┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼
5400 5150 4900 4650 4400 4150 3900 3650 3400 3150 2900 2650 2400億年前
5.3880億年前 4.8540億年前 4.43億年前4.1920億年前 3.5890億年前 2.9890億年前
カンブリア紀 | オルドビス紀|シルル紀| デボン紀 | 石炭紀 | ペルム紀
万年 5340 4200 2420 6030 6000 4700万年┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼
5400 5150 4900 4650 4400 4150 3900 3650 3400 3150 2900 2650 2400億年前
←────────────────────-中生代─────────────────────→
2.51902億年前 2.0130億年前 1.4500億年前
┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼
25000 23500 22000 20500 19000 17500 16000 14500 13000 11500 10000 8500 6000万年前
2.51902億年前 2.0130億年前 1.4500億年前
三畳紀 | ジュラ紀 | 白亜紀
万年 5060.2 5640 7900 万年┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼───┼
25000 23500 22000 20500 19000 17500 16000 14500 13000 11500 10000 8500 6000万年前
←──────-新生代-──────→
6600万年前 2303万年前 258万年前
┼──┼──┼──┼──┼──┼──┼
6600 5500 4400 3300 2200 1100 0万年前
6600万年前 2303万年前 258万年前
古第三紀 | 新第三紀 | ←第四紀
万年 4297 2045 258万年┼──┼──┼──┼──┼──┼──┼
6600 5500 4400 3300 2200 1100 0万年前
←────────-古第三紀─────────→|←───新第三紀───→|第四紀 |
6600万年前 5600万年前 3390万年前 2303万年前 533万3000年前
┼────┼────┼────┼────┼────┼────┼────┼────┼
6600 5900 5200 3500 2800 2100 1400 700 0万年前
鮮新世533万3000年前 275万3000年
更新世 258万年前 256万8300年
完新世 1万1700年前 1万1700年
6600万年前 5600万年前 3390万年前 2303万年前 533万3000年前
暁新世 | 始新世 | 漸新世 | 中新世 | | |←鮮新世|更新世|完新世
万年1000 2210 1087 1769.7 275.3 ┼────┼────┼────┼────┼────┼────┼────┼────┼
6600 5900 5200 3500 2800 2100 1400 700 0万年前
鮮新世533万3000年前 275万3000年
更新世 258万年前 256万8300年
完新世 1万1700年前 1万1700年
TOP|
地球史円グラフ(年数割合)
|TOP
新生代1%
先カンブリア時代:41億2820万年(88%) カンブリア紀:5340万年─┐
顕生代 :5億3880万年(12%) 古中 オルドビス紀:4200万年 │
生生冥王代 シルル紀 :2420万年 古生代2億8690万年
古第三紀 暁新世:1000万年 代代 13% デボン紀 :6030万年 │
始新世:2210万年 6%4% 石炭紀 :6000万年 │
漸新世:1087万年 ペルム紀 :4700万年─┘
原生代 太古代 三畳紀 :5060万年─┐
新第三紀 中新世:1769万7000年 42% 34% ジュラ紀 :5630万年 中生代1億8590万年
鮮新世: 275万3000年 白亜紀 :7900万年─┘
古第三紀 :4297万年─┐
第四紀 更新世: 256万8300年 新第三紀 :2045万年 新生代 6600万年
完新世: 1万1700年 第四紀 : 258万年─┘
先カンブリア時代:41億2820万年(88%) カンブリア紀:5340万年─┐
顕生代 :5億3880万年(12%) 古中 オルドビス紀:4200万年 │
生生冥王代 シルル紀 :2420万年 古生代2億8690万年
古第三紀 暁新世:1000万年 代代 13% デボン紀 :6030万年 │
始新世:2210万年 6%4% 石炭紀 :6000万年 │
漸新世:1087万年 ペルム紀 :4700万年─┘
原生代 太古代 三畳紀 :5060万年─┐
新第三紀 中新世:1769万7000年 42% 34% ジュラ紀 :5630万年 中生代1億8590万年
鮮新世: 275万3000年 白亜紀 :7900万年─┘
古第三紀 :4297万年─┐
第四紀 更新世: 256万8300年 新第三紀 :2045万年 新生代 6600万年
完新世: 1万1700年 第四紀 : 258万年─┘
TOP|
地球史時計(時:分:秒)
|TOPTOP|
地球史カレンダー(平年月日)
|TOP開始時刻 開始月日 開始年代 地質時代 出来事 年数
00:00:00 1月 1日 45億6700万年前 冥王代 地球誕生 月誕生 5億3600万年 ↑
03:07:50 2月18日 40億3100万年前 太古代 生命誕生 15億年 先カンブリア時代
10:57:23 6月16日 25億年前 原生代 光合成生物の進化 19億5900万年 ____↓____
21:10:39 11月18日 5億3880万年前 古生代 カンブリア大爆発 2億7690万年 ↑
22:41:09 12月12日 2億5190万年前 中生代 恐竜の時代 1億8590万年 顕生代
23:39:20 12月27日 6600万年前 新生代 霊長類の出現 6600万年 ____↓____
23:59:04 12月31日 30万年前 (現生人類) ホモサピエンス出現 新生代中
23:59:59 12月31日 6000年前 (有史時代) 文字の発明 新生代中
年数→時間数:(億年数÷46)×24時間と余りを分秒計算
開始年代→開始時刻=((46-億年前)/46)×24と余り分秒計算、または、時間数→時間数累計(時分秒)
年数→日数:(億年数÷46)×365日
開始年代→開始月日=日数→通日(日数累計+1)から月日変換
※年代は国際層序委員会,2023年09月によります
TOP|

地球上の生物種数は推定約870万とする研究報告発表(2011) … 未知の生物を含む
陸上種650万、海洋種220万 → 陸上種の86%、海洋種の91%が未知種
国際自然保護連合はレッドリストを公開(2019) … 絶滅危惧種の数は28,338種
1975年~2000年の25年間に年平均4万種、13分間に1種の生き物が絶滅
環境省レッドリスト(日本の生物)
野生動物の3割が絶滅危惧種
野生植物の2割が絶滅危惧種
F:力、q:(±)電荷量、m:質量、r:距離
クーロンの法則の式は万有引力の法則の式に似ているが、万有引力には斥力がない
反陽子の発見→クォークの存在予言と発見→ヒッグス粒子の発見
電子以外の素粒子は20世紀に発見、ヒッグス粒子は2012年に発見
アルファ線、陽子線、ベータ線、宇宙からのミュー粒子:物質中の電子の電気力で減速して止まる
ガンマ線:物質中の電子に何回か衝突すると吸収されて消滅する(一部又は全部が電子のエネルギーになる)
光子は電子に衝突して、散乱、吸収、電子・陽電子対に変換される
中性子線:物質中の原子核に衝突して吸収されて消滅する
元素記号=【赤:常温で気体(H,N,O,F,Cl,18族)、青:常温で液体(Hg,Br)、黒:常温で固体】
背景の色=非金属、アルカリ金属、アルカリ土類金属、遷移金属、ハロゲン、貴ガス、典型金属
金属元素:金属元素、非金属元素:金属元素以外、半金属元素:金属と非金属との境界付近の元素(両方の性質)
遷移元素(横の類似性):3~12族、3族のうち原子番号=57~70と原子番号=89~102は内部遷移元素
典型元素(縦の類似性):遷移元素以外、典型元素には非金属元素と金属元素がある、テネシンも典型元素に含む
内部遷移元素:原子番号=57~70:ランタノイド(La~Yb)、原子番号=89~102:アクチノイド(Ac~No)
希土類元素(17元素):Scスカンジウム,Yイットリウム,ランタノイド=(「ランタン」+「もどき」)
貴ガスは量的に少ないので希ガスとも言われる化合反応を起こしにくい不活性ガス(最外殻までの電子が満杯状態)
2005年に英語表記がrare gasからnoble gasに改められ,日本語での表記も希ガスから貴ガスへと変更になりました
令和3年度の教科書より、日本化学会の提言も踏まえて高貴なガスという意味から貴ガスと表記を変更しています
化学反応しやすい:電子が満杯+1個余計(Na+)、電子が満杯-1個少ない(Cl-)=化学反応を起こしてNaClになる
第12族から第16族は先頭元素名で呼ばれる(亜鉛族,ホウ素族,炭素族,窒素族,酸素族)
ニクトゲン:15族に位置する5つの元素(N,P,As,Sb,Bi),カルコゲン:16族に位置する5つの元素(O,S,Se,Te,Po)
人工元素:天然に存在する元素は原則、原子番号92Uウランまでですが、原子番号92までのうち4つの例外元素
=43Tcテクネチウム,61Pmプロメチウム,85Atアスタチン,87Frフランシウム(同位体)および原子番号が93以上の
超ウラン元素(アクチノイドの一部93~103,104-118の24個),いずれも放射性元素
→┼→┼──→┼──→┼────→┼────→┼──────→┼────→┼──→┼→
殻 K L L M M N M N O N O P N O P Q O P Q P Q Q
軌道1s→2s→2p→3s→3p→4s→3d→4p→5s→4d→5p→6s→4f→5d→6p→7s→5f→6d→7p→6f→7d→7f
(E)1s<2s<2p<3s<3p<4s<3d<4p<5s<4d<5p<6s<4f<5d<6p<7s<5f<6d<7p<6f<7d<7f
という順で詰まっていきます(銅などいくつかの例外もある)
○主量子数:n = 1,2,3,…,7 (正の整数) = 電子殻:K,L,M,N,…殻
○主量子数nと方位量子数l(エル)との関係…0≦l≦n-1
○方位量子数l(エル)と磁気量子数mlとの関係…-l≦ml≦l
エネルギー準位(E)は、通常、主量子数(n)が大きくなるほど高く、主量子数(n)が同じ値の場合には、
方位量子数(l:エル)の値が大きくなるほど高いが、n≧3の場合は、高低の逆転が生じる(4s<3d)
このように見ると、M殻の3d軌道の前にN殻の4s軌道が詰まるなど「K殻から順に」のルールが崩れています
実はこれこそが「遷移元素」が存在する理由です
電子軌道は、エネルギーの低い順に収容される(4s<3d)
例として原子番号21、電子数21のスカンジウムを見ていきます
カルシウムは電子数20ですから、電子1個を残して全てカルシウムと同じように詰まります
カルシウムはK殻に2個、L殻に8個、M殻に8個入ると、あと10個の電子がM殻に入れるのですが、
残り2個の電子は、M殻より軌道エネルギーの低いN殻に先に入ります(4s<3d)
(Ca20の場合、K2,L8,M10でなく、K2,L8,M8,N2)
K L L M M N
20Ca:1s2→2s2→2p6→3s2→3p6→4s2
電子数 2 4 10 12 18 20
アップスピン↑とダウンスピン↓が1つずつ入る(1つの軌道には電子は上下逆さのスピンで入る)電子対
同じエネルギーの軌道にはバラバラに、スピンを揃えて入るという規則があります(フントの規則)不対電子
そしてスカンジウムの最後の1個は、N殻4s軌道の次にエネルギーの低いM殻3d軌道に詰まります
K L L M M N M
21Sc:1s2→2s2→2p6→3s2→3p6→4s2→3d1
電子数 2 4 10 12 18 20 21
この先最外殻電子は4s軌道の2個のまま、3d軌道に入っていくことになり、これが「遷移元素」が生じる原因です
亜鉛族元素:第12族元素は、d軌道が閉殻状態ゆえ典型元素的性質を持つため、以前は「典型元素」に分類され
ていましたが、IUPACの最近の勧告にあわせて、日本も2022年度からは「遷移元素」に含めるとしています
従来の周期表の中には、(La~Lu)及び((Ac~Lr)の各15元素を「内部遷移元素」としているものもありますが、
電子配置からするとLu及びLrは「内部遷移元素」でなく「遷移元素」です
本来、内部遷移元素は(n-2)f1-14の電子を持ち、各シリーズ14個の元素からなることが特長です
ランタノイド(La~Lu)15個のうちLuを除外した(La~Yb)が内部遷移元素14個、Luは遷移元素
(La~Yb)+(Lu~Hg):(14+10)元素=4f+5d
アクチノイド(Ac~Lr)15個のうちLrを除外した(Ac~No)が内部遷移元素14個、Lrは遷移元素
(Ac~No)+(Lr~Cn):(14+10)元素=5f+6d
1番外側の電子殻に収容されている電子のことを最外殻電子といいます
貴ガスの最外殻電子はヘリウムだけ2個で、他の貴ガスはすべて最外殻電子は8個で安定な電子配置をしています
このように最大数の電子が収容された電子殻を閉殻といいます
化学結合や化学的性質に関与する電子のことを価電子といいます
最外殻電子がちょうどその電子殻の最大収容数の場合、または最外殻電子が8個の場合、価電子の数は0とする
貴ガスの場合は最外殻までの電子が満杯状態で、化学反応をそもそも起こさないので、価電子の数はゼロです
価電子の定義は曖昧です(貴ガスは反応性が低いから価電子数はすべて0としますと言う程度の定義です!)
同一元素の原子はその原子核に必ず同数の陽子を持っているが、中性子数の異なる原子核(核種)を同位体と呼ぶ
同位体は陽子と軌道電子の数が等しいため化学的性質はほぼ等しいが、物理的性質は異なる
同位体のうち放射性を有するものを放射性同位体、放射性を有しないものを安定同位体と呼ぶ
背景の色=
古代、
13世紀、
15世紀、
17世紀、
18世紀、
19世紀、
20世紀、
21世紀
元素数 = 10個、 1個、 1個、 1個、 21個、 49個、 30個、 5個
地表露出:有用資源の採掘
熱分解・電気分解:化合物を単体に分解
分光器:分光分析、スペクトル分析
周期律の発見:三つ組元素、未知元素の予言(周期表の空欄)
放射線:放射性元素の発見
加速器:原子核反応、人工元素、現代の錬金術
スペクトルの特徴=s軌道(sharp)、p軌道(principal)、d軌道(diffuse)、f軌道(fundamental)
背景の色=非金属、アルカリ金属、アルカリ土類金属、遷移金属、ハロゲン、貴ガス、典型金属
遷移元素:3~12族、3族のうち原子番号=57~70と原子番号=89~102は内部遷移元素
[貴ガスの元素記号]:電子配置の共通項を簡略形式で表現、例:Se=[Ar],3d10,4s2,4p4
クロム(Cr)と銅(Cu)の電子配置は次の通りです
最外殻電子4sから電子を一つ奪って3d軌道に入っています
●クロム(Cr,24):1s2,2s2,2p6,3s2,3p6,3d5,4s1
d軌道に5個の不対電子が入ることで、スピンが同じ方向を向きエネルギー的に安定化します
(d軌道が半閉殻)
●銅(Cu,29) :1s2,2s2,2p6,3s2,3p6,3d10,4s1
d軌道に10個の電子対が入ることで、d軌道の電子が全て互いに逆向きスピンで充填され、
エネルギー的に安定化します(d軌道が閉殻)
パラジウム(Pd)の電子配置は次の通りです
最外殻電子5sから電子を二つ奪って4d軌道に入っています
●パラジウム(Pd,46):1s2,2s2,2p6,3s2,3p6,3d10,4s2,4p6,4d10,5s0
先に5s軌道から電子が入り始めますが、4d軌道に電子が入り始め4d軌道の電子が増え、
Pdでは5s(↑↓)4d(↑↓)(↑↓)(↑↓)(↑)(↑)ではなく
5s()4d(↑↓)(↑↓)(↑↓)(↑↓)(↑↓)という電子配置になります(d軌道が閉殻)
遷移金属はエネルギーの近い軌道(dとs,dとf)を持つため、順番が変わることが多いので注意が必要です
電子はスピンの向きを揃えたがる性質と、できるだけ離れた電子軌道に入ろうとする二つの重要な性質
があり、最終的に原子全体としていかにバランスよく電子配置するかという観点で電子を配置します
dブロック(3d~6d)の例外1:
d軌道の副殻は、s軌道の副殻から1個の電子(パラジウムの場合は2個)を「借ります」
fブロック(4f~5f)の例外2:
d軌道の副殻は、f軌道の副殻から1個の電子(トリウムの場合は2個)を「借りる」ことがよくあります
pブロック(2p~7p)の例外3:
1970年、ローレンシウム(Lr,103)の基底状態の電子配置は、構造原理に従って、[Rn]5f146d17s2であり、
同族体であるルテチウムの[Xe]4f145d16s2とも合致すると予測された
しかし翌年、この予測に疑義を唱え、その代わり、[Rn]5f147s27p1という異常な電子配置を取るとする
計算結果が公表された
相対性理論はアインシュタイン1人がほぼ独力でつくりあげた
量子論は何人もの天才物理学者がアイデアを出し合って作られた
注:反射と透過の重ね合わせ=粒子の存在場所の重ね合わせ、(広がりがある)波ならではの現象
TOP|
地球史年表(等間隔)
|TOP
←─────────-先カンブリア時代─────────→←───────────-顕生代-───────────→
←──────────────先カンブリア時代──────────────→
←─────────────────────────-顕生代─────────────────────────→
←─────────────-新生代──────────────→
| 冥王代 | 太古代 | 原生代 | 古生代 | 中生代 | 新生代 |
| 45億6700万年前 | 40.31 | 25 | 5.3880 | 2.51902 | 0.6600 |
←──────────────先カンブリア時代──────────────→
| 冥王代 | 太古代 | 原生代 | |||||
| 原太古代 | 古太古代 | 中太古代 | 新太古代 | 古原生代 | 中原生代 | 新原生代 | |
| 45.67 | 40.31 | 36 | 32 | 28 | 25 | 16 | 10 |
←─────────────────────────-顕生代─────────────────────────→
| 古生代 | 中生代 | 新生代 | |||||||||
| カンブリア紀 | オルドビス紀 | シルル紀 | デボン紀 | 石炭紀 | ペルム紀 | 三畳紀 | ジュラ紀 | 白亜紀 | 古第三紀 | 新第三紀 | 第四紀 |
| 5.3880 | 4.8540 | 4.4300 | 4.1920 | 3.5890 | 2.9890 | 2.51902 | 2.0130 | 1.4500 | 0.6600 | 0.2303 | 0.0258 |
←─────────────-新生代──────────────→
| 古第三紀 | 新第三紀 | 第四紀 | ||||
| 暁新世 | 始新世 | 漸新世 | 中新世 | 鮮新世 | 更新世 | 完新世 |
| 0.6600 | 0.5600 | 0.3390 | 0.2303 | 0.05333 | 0.0258 | 0.000117 |
| 全生物の共通祖先(コモノート) |
|---|
|
┌───非核酸複製生物(絶滅) │ 化学進化…プロテノイドミセル─┤ ┌─低温菌(絶滅) (有機物) アミノ酸の │ │ 熱重合体の │ ├─常温菌(絶滅) 分子集合体 │ │ (生命の起源) │ ┌─DNAゲノム生物─┤ ┌───真正細菌(バクテリア) │ │ │ │ │ │ ├─超好熱菌─┴─┬─古細菌 (アーキア) │ │ │(共通祖先) │ └─┤ │ └─真核生物(ユーカリア) │ │ │ └─好熱菌(絶滅) │ └─RNAゲノム生物(絶滅)
プロテノイド・ミクロスフェア … プロテノイド:蛋白質類似体、
ミクロスフェア:ミクロンサイズの球状粒子、 ミセル:分子集合体 LUCA「Last Universal Common Ancestor」(最終普遍共通祖先)は、 その起源を約40億年前にまでさかのぼることのできる単細胞生物である |
| 3ドメイン説 |
|
┌───真正細菌(原核生物) │ 起源生物─┴─┬─古細菌 (原核生物) (共通祖先) │ └─真核生物──原生生物、植物、菌類、動物 ※ドメイン:界より上位の階級、古細菌の幹からの分岐は、最初が真正細菌で次が真核生物という説が主流 |
| 五界説 |
|
┌───────原核生物 … 核膜がなく、核酸が細胞質に直接露出している │ 起源生物─┤ ┌─原生生物 … 真核生物のうち、菌界にも植物界にも動物界にも属さない生物の総称 (共通祖先)│ │ └真核生物─┼─植物 … 細胞壁を有し、独立栄養で光合成を行うことができる生物をいう │ ├─菌類 … 細胞壁をもつが、専ら動植物に寄生し、生活の全てを宿主に依存している │ └─動物 … 細胞壁をもたず、従属栄養で栄養源は植物に依存している ※真核生物:真核とよばれる核膜で囲まれた明確な細胞核をもっている生物の総称 |
┌──────────────────────────────────────────────────────┐
│ 起源生物 │
│ ┏━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┓ │
│ ┃ 原核生物 ┃ │
│ ┃ ┌─────────────────┐┌────────────┐ ┃ │
│ ┃ │ 真正細菌(バクテリア) ││ 古細菌(アーキア) │ ┃ │
│ ┃ │シアノバクテリア(藍藻、藍色細菌)││メタン生成菌、超好熱菌 │ ┃ │
│ ┃ │紅色硫黄細菌、亜硫酸菌、枯草菌 ││高度酸性菌、高度好塩菌 │ ┃ │
│ ┃ │大腸菌、乳酸菌、放線菌 │└────────────┘ ┃ │
│ ┃ └─────────────────┘ 原核生物:核膜がない ┃ │
│ ┃ 単細胞生物 真核生物:核膜がある ┃ │
│ ┗━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┛ │
│┌────────────────────────────────────────────────────┐│
││ 真核生物 ││
││┏━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┓││
││┃ 原生生物 ┃││
││┃┌────────────────────────────────────────────────┐┃││
││┃│ 植物性 │┃││
││┃│ ┌─────────────────────────┐ │┃││
││┃│ │ 藻類 │ │┃││
││┃│ │┌────────┐┌────┐┌───────┐│ │┃││
││┃│ ││ 緑藻類 ││紅藻類 ││ 褐藻類 ││ │┃││
││┃│ ││アオノリ、アオサ││テングサ││コンブ、ワカメ││ │┃││
││┃│ │└────────┘└────┘└───────┘│ │┃││
││┃│ └─────────────────────────┘ │┃││
││┃│┌──────┐┌──────┐┌──────┐┌────┐┌───────┐┌───────┐│┃││
││┃││ミドリムシ類││ 渦鞭毛藻類 ││ 珪藻類 ││ 卵菌類 ││ 変形菌類 ││ 細胞性粘菌類 ││┃││
││┃││ミドリムシ ││ ツノモ ││ハネケイソウ││ミズカビ││ムラサキホコリ││タマホコリカビ││┃││
││┃│└──────┘│ヤコウチュウ│└──────┘└────┘└───────┘└───────┘│┃││
││┃│ ↑ └──────┘ ↑ │┃││
││┃│ │ └真正粘菌 │┃││
││┃│ └鞭毛運動をする動物的性質を持ちながら、同時に植物として葉緑体を持ち光合成を行う「単細胞生物」│┃││
││┃│ │┃││
││┃│ クロロフィルa+b:ミドリムシ類、緑藻類、植物 │┃││
││┃│ クロロフィルa :紅藻類 │┃││
││┃│ クロロフィルa+c:渦鞭毛藻類、珪藻類、褐藻類 │┃││
││┃└────────────────────────────────────────────────┘┃││
││┃ ┌───────────────────────────────┐ ┃││
││┃ │ 動物性 │ ┃││
││┃ │┌────┐┌───────┐┌─────┐┌───────┐│ ┃││
││┃ ││肉質虫類││ 鞭毛虫類 ││ 繊毛虫類 ││ 胞子虫類 ││ ┃││
││┃ ││アメーバ││トリパノソーマ││ゾウリムシ││マラリア病原虫││ ┃││
││┃ │└────┘└───────┘└─────┘└───────┘│ ┃││
││┃ │ 単細胞生物 │ ┃││
││┃ └───────────────────────────────┘ ┃││
││┃ 藻類:酸素発生型光合成生物で地上生息以外の総称 ┃││
││┃ 水生・海生生息 ┃││
││┃ バクテリアに属する藻類(単細胞生物) ┃││
││┃ シアノバクテリア、藍藻、藍色細菌 ┃││
││┃ 真核生物に属する藻類(単細胞生物、多細胞生物) ┃││
││┃ ┃││
││┃ 菌類:ほとんどが多細胞生物、酵母は単細胞生物 ┃││
││┃ 地衣類は菌類と藻類の共生生物 ┃││
││┗━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┛││
││┏━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┓││
││┃ 植物 ┃││
││┃ ┌─────────────────────┐ ┃││
││┃ │ コケ植物 │ ┃││
││┃ │┌─────┐┌─────┐┌─────┐│ ┃││
││┃ ││ コケ類 ││ツノゴケ類││ 鮮類 ││ ┃││
││┃ ││ゼニゴケ ││ツノゴケ ││ミズゴケ ││ ┃││
││┃ │└─────┘└─────┘└─────┘│ ┃││
││┃ └─────────────────────┘ ┃││
││┃ ┌──────────────────────────────────────────────┐ ┃││
││┃ │ 維管束植物 │ ┃││
││┃ │ ┌──────────────────────────────┐ │ ┃││
││┃ │ │ シダ植物 │ │ ┃││
││┃ │ │┌─────┐┌────┐┌──────┐┌───────┐│ │ ┃││
││┃ │ ││ シダ類 ││トクサ類││マツバラン類││ヒゲノカズラ類││ │ ┃││
││┃ │ ││ゼンマイ ││トクサ ││マツバラン ││ヒゲノカズラ ││ │ ┃││
││┃ │ │└─────┘└────┘└──────┘└───────┘│ │ ┃││
││┃ │ └──────────────────────────────┘ │ ┃││
││┃ │┌────────────────────────────────────────────┐│ ┃││
││┃ ││ 種子植物 ││ ┃││
││┃ ││┌──────────────────────┐┌──────────────────┐││ ┃││
││┃ │││ 裸子植物 ││ 被子植物 │││ ┃││
││┃ │││┌──────┐┌─────┐┌─────┐││┌──────┐┌────────┐│││ ┃││
││┃ ││││ 球果類 ││ ソテツ類 ││イチョウ類││││ 単子葉類 ││ 双子葉類 ││││ ┃││
││┃ ││││スギ、ヒノキ││ ソテツ ││イチョウ ││││イネ、コムギ││サクラ、アサガオ││││ ┃││
││┃ │││└──────┘└─────┘└─────┘││└──────┘└────────┘│││ ┃││
││┃ ││└──────────────────────┘└──────────────────┘││ ┃││
││┃ │└────────────────────────────────────────────┘│ ┃││
││┃ │ 種子で増える:裸子植物・被子植物 │ ┃││
││┃ └──────────────────────────────────────────────┘ ┃││
││┃ 胞子で増える:藻類・菌類・コケ植物・シダ植物、維管束なし=コケ植物、維管束あり=シダ植物 ┃││
││┗━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┛││
││┏━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┓││
││┃ 菌類 ┃││
││┃ ┌──────┐┌─────┐┌──────┐┌──────┐┌───────┐ ┃││
││┃ │ 担子菌類 ││不完全菌類││ 子嚢菌類 ││ 接合菌類 ││ ツボカビ類 │ ┃││
││┃ │ シイタケ ││アオカビ ││アカパンカビ││クモノスカビ││カエルツボカビ│ ┃││
││┃ └──────┘└─────┘└──────┘└──────┘└───────┘ ┃││
││┃ ツボカビ類:鞭毛あり ┃││
││┃ 他 :鞭毛なし ┃││
││┃ カビの姿=菌糸+胞子 ┃││
││┃ キノコ=子実体+菌糸体+胞子 ┃││
││┗━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┛││
││┏━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┓││
││┃ 動物 ┃││
││┃ ┌────────────────┐ ┃││
││┃ │ 無胚葉 │ ┃││
││┃ │┌──────────────┐│ ┃││
││┃ ││ 海綿動物 ││ ┃││
││┃ ││カイロオウドウケツ、カイメン││ ┃││
││┃ │└──────────────┘│ ┃││
││┃ └────────────────┘ ┃││
││┃ ┌──────────────────────────────┐ ┃││
││┃ │ 二胚葉 │ ┃││
││┃ │ ┌─────┐┌───────────┐ │ ┃││
││┃ │ │ 有櫛動物 ││ 刺胞動物 │ │有櫛動物:刺胞なし┃││
││┃ │ │クシクラゲ││サンゴ、イソギンチャク│ │刺胞動物:刺胞あり┃││
││┃ │ └─────┘└───────────┘ │ ┃││
││┃ │ ┌───────────┐ │ ┃││
││┃ │ │ 平板動物 │ │ ┃││
││┃ │ │ センモウヒラムシ │ │ ┃││
││┃ │ └───────────┘ │ ┃││
││┃ └──────────────────────────────┘ ┃││
││┃ ┌──────────────────────────────────────────┐ ┃││
││┃ │ 三胚葉 │ ┃││
││┃ │┌────────────────────────────────────────┐│ ┃││
││┃ ││ 旧口動物 ││ ┃││
││┃ ││ ┌──────────────────────────────────┐ ││ ┃││
││┃ ││ │ 無体腔 │ ││ ┃││
││┃ ││ │┌────────────────────────────────┐│ ││ ┃││
││┃ ││ ││ 扁形動物 ││ ││ ┃││
││┃ ││ ││┌──────┐┌──────┐┌──────┐┌──────┐││ ││ ┃││
││┃ ││ │││ 渦虫類 ││ 吸虫類 ││ 単生類 ││ 条虫類 │││ ││ ┃││
││┃ ││ │││プラナリア ││ ジストマ ││フタゴムシ ││サナダムシ │││ ││ ┃││
││┃ ││ ││└──────┘└──────┘└──────┘└──────┘││ ││ ┃││
││┃ ││ │└────────────────────────────────┘│ ││ ┃││
││┃ ││ └──────────────────────────────────┘ ││ ┃││
││┃ ││ ┌────────────────────┐ ││ ┃││
││┃ ││ │ 偽体腔 │ ││ ┃││
││┃ ││ │┌────────┐┌────────┐│ ││ ┃││
││┃ ││ ││ 冠輪動物 ││ 脱皮動物 ││ ││ ┃││
││┃ ││ ││┌──────┐││┌──────┐││ ││ ┃││
││┃ ││ │││ 輪形動物 ││││ 線形動物 │││ ││ ┃││
││┃ ││ │││ツボワムシ ││││カイチュウ │││ ││ ┃││
││┃ ││ │││ネズミワムシ││││線虫、鞭虫 │││ ││ ┃││
││┃ ││ ││└──────┘││└──────┘││ ││ ┃││
││┃ ││ │└────────┘└────────┘│ ││ ┃││
││┃ ││ └────────────────────┘ ││ ┃││
││┃ ││┌──────────────────────────────────────┐││ ┃││
││┃ │││ 真体腔 │││ ┃││
││┃ │││┌────────────┐┌──────────────────────┐│││ ┃││
││┃ ││││ 冠輪動物 ││ 脱皮動物 ││││ ┃││
││┃ ││││ ┌──────┐ ││┌────────────────────┐││││ ┃││
││┃ ││││ │ 環形動物 │ │││ 節足動物 │││││ ┃││
││┃ ││││ │ヒル、ミミズ│ │││ ┌────────────┐ │││││ ┃││
││┃ ││││ └──────┘ │││ │ 鋏角類 │ │││││ ┃││
││┃ ││││┌──────────┐│││ │クモ、サソリ、カブトガニ│ │││││ ┃││
││┃ │││││ 軟体動物 ││││ └────────────┘ │││││ ┃││
││┃ │││││┌────────┐││││ ┌────────────┐ │││││ ┃││
││┃ ││││││ 二枚貝類 │││││ │ 多足類 │ │││││ ┃││
││┃ ││││││ハマグリ、アサリ│││││ │ ムカデ、ゲジ、ヤスデ │ │││││ ┃││
││┃ │││││└────────┘││││ └────────────┘ │││││ ┃││
││┃ │││││┌────────┐││││ ┌────────────────┐ │││││ ┃││
││┃ ││││││ 巻貝類 │││││ │ 甲殻類 │ │││││ ┃││
││┃ ││││││アワビ、サザエ │││││ │ エビ、カニ、ミジンコ、フジツボ │ │││││ ┃││
││┃ ││││││カタツムリ │││││ └────────────────┘ │││││ ┃││
││┃ │││││└────────┘││││┌──────────────────┐│││││ ┃││
││┃ │││││┌────────┐│││││ 六脚類 ││││││ ┃││
││┃ ││││││ 頭足類 ││││││┌────────────────┐││││││ ┃││
││┃ ││││││ タコ、イカ │││││││ 内顎類 │││││││ ┃││
││┃ ││││││ オウムガイ │││││││ トビムシ、コムシ、カマアシムシ │││││││ ┃││
││┃ │││││└────────┘│││││└────────────────┘││││││ ┃││
││┃ ││││└──────────┘││││ ┌──────────────┐ ││││││ ┃││
││┃ │││└────────────┘│││ │ 昆虫類 │ ││││││ ┃││
││┃ │││ │││ │カブトムシ、ゴミムシ │ ││││││ ┃││
││┃ │││ │││ │チョウ、ガ、トビケラ │ ││││││ ┃││
││┃ │││ │││ │ハエ、カ、アブ、ノミ、シラミ│ ││││││ ┃││
││┃ │││ │││ │スズメバチ、ミツバチ、アリ │ ││││││ ┃││
││┃ │││ │││ │セミ、カメムシ、アザミウマ │ ││││││ ┃││
││┃ │││ │││ │バッタ、キリギリス、コオロギ│ ││││││ ┃││
││┃ │││ │││ │カマキリ、カゲロウ、トンボ │ ││││││ ┃││
││┃ │││ │││ │ゴキブリ、シロアリ │ ││││││ ┃││
││┃ │││ │││ └──────────────┘ ││││││ ┃││
││┃ │││ ││└──────────────────┘│││││ ┃││
││┃ │││ │└────────────────────┘││││ ┃││
││┃ │││ └──────────────────────┘│││ ┃││
││┃ ││└──────────────────────────────────────┘││ ┃││
││┃ │└────────────────────────────────────────┘│ ┃││
││┃ │┌────────────────────────────────────────┐│ ┃││
││┃ ││ 新口動物 ││ ┃││
││┃ ││ ┌────┐┌───────────────┐ ││ ┃││
││┃ ││ │毛顎動物││ 棘皮動物 │ ││ ┃││
││┃ ││ │ヤムシ ││ウミユリ、ヒトデ、ウニ、ナマコ│ ││ ┃││
││┃ ││ └────┘└───────────────┘ ││ ┃││
││┃ ││┌──────────────────────────────────────┐││ ┃││
││┃ │││ 脊索動物 │││ ┃││
││┃ │││ ┌──────────────────┐ │││ ┃││
││┃ │││ │ 原索動物 │ │││ ┃││
││┃ │││ │┌───────┐┌───────┐│ │││ ┃││
││┃ │││ ││ 頭索類 ││ 尾索類 ││ │││ ┃││
││┃ │││ ││ ナメクジウオ ││ ホヤ ││ │││ ┃││
││┃ │││ │└───────┘└───────┘│ │││ ┃││
││┃ │││ └──────────────────┘ │││ ┃││
││┃ │││┌────────────────────────────────────┐│││ ┃││
││┃ ││││ 脊椎動物 ││││ ┃││
││┃ ││││ ┌──────┐ ││││ ┃││
││┃ ││││ │ 無顎類 │ ││││ ┃││
││┃ ││││ │ヤツメウナギ│ ││││ ┃││
││┃ ││││ └──────┘ ││││ ┃││
││┃ ││││┌──────────────────────────────────┐││││ ┃││
││┃ │││││ 顎口類 │││││ ┃││
││┃ │││││ ┌──────┐ │││││ ┃││
││┃ │││││ │ 軟骨魚類 │ │││││ ┃││
││┃ │││││ │サメ、エイ │ │││││ ┃││
││┃ │││││ └──────┘ │││││ ┃││
││┃ │││││┌────────────────────────────────┐│││││ ┃││
││┃ ││││││ 硬骨魚類 ││││││ ┃││
││┃ ││││││ ┌──────┐ ││││││ ┃││
││┃ ││││││ │ 条鰭類 │ ││││││ ┃││
││┃ ││││││ │コイ、タイ │ ││││││ ┃││
││┃ ││││││ └──────┘ ││││││ ┃││
││┃ ││││││┌──────────────────────────────┐││││││ ┃││
││┃ │││││││ 肉鰭類 │││││││ ┃││
││┃ │││││││ ┌───────┐┌─────┐ │││││││ ┃││
││┃ │││││││ │シーラカンス類││ 肺魚類 │ │││││││ ┃││
││┃ │││││││ │ラティメリア ││ハイギョ │ │││││││ ┃││
││┃ │││││││ └───────┘└─────┘ │││││││ ┃││
││┃ │││││││┌────────────────────────────┐│││││││ ┃││
││┃ ││││││││ 四足類 ││││││││ ┃││
││┃ ││││││││┌───┐┌─────────────────────┐││││││││ ┃││
││┃ │││││││││両生類││ 羊膜類 │││││││││ ┃││
││┃ │││││││││カエル││┌─────┐┌─────┐┌─────┐│││││││││ ┃││
││┃ │││││││││イモリ│││ 爬虫類 ││ 鳥類 ││ 哺乳類 ││││││││││ ┃││
││┃ ││││││││└───┘││ヘビ、ワニ││ハト、タカ││ヒト、イヌ││││││││││ ┃││
││┃ ││││││││ ││トカゲ ││スズメ ││クジラ ││││││││││ ┃││
││┃ ││││││││ │└─────┘└─────┘└─────┘│││││││││ ┃││
││┃ ││││││││ └─────────────────────┘││││││││ ┃││
││┃ │││││││└────────────────────────────┘│││││││ ┃││
││┃ ││││││└──────────────────────────────┘││││││ ┃││
││┃ │││││└────────────────────────────────┘│││││ ┃││
││┃ ││││└──────────────────────────────────┘││││ ┃││
││┃ │││└────────────────────────────────────┘│││ ┃││
││┃ ││└──────────────────────────────────────┘││ ┃││
││┃ │└────────────────────────────────────────┘│ ┃││
││┃ │ │ ┃││
││┃ │ 旧口動物・新口動物 │ ┃││
││┃ │ 体壁と消化管の隙間を体腔と呼び、体腔が内胚葉で包まれているものを真体腔と呼ぶ │ ┃││
││┃ │ 初期胚の体内を貫通する腸管が貫入した入口(先にできた口なので原口とか前口と呼ぶ)と │ ┃││
││┃ │ 初期胚の体内を貫通した腸管が反対側に出た出口(後にできた口なので後口と呼ぶ)が各々 │ ┃││
││┃ │ 何になるかによって旧口(前口)か新口(後口)かに分ける │ ┃││
││┃ │ │ ┃││
││┃ │ 入口が口になり、出口が肛門になるのが旧口(前口) … 入口:旧い口 │ ┃││
││┃ │ 出口が口になり、入口が肛門になるのが新口(後口) … 出口:新しい口 │ ┃││
││┃ │ │ ┃││
││┃ └──────────────────────────────────────────┘ ┃││
││┗━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┛││
│└────────────────────────────────────────────────────┘│
└──────────────────────────────────────────────────────┘
│ 起源生物 │
│ ┏━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┓ │
│ ┃ 原核生物 ┃ │
│ ┃ ┌─────────────────┐┌────────────┐ ┃ │
│ ┃ │ 真正細菌(バクテリア) ││ 古細菌(アーキア) │ ┃ │
│ ┃ │シアノバクテリア(藍藻、藍色細菌)││メタン生成菌、超好熱菌 │ ┃ │
│ ┃ │紅色硫黄細菌、亜硫酸菌、枯草菌 ││高度酸性菌、高度好塩菌 │ ┃ │
│ ┃ │大腸菌、乳酸菌、放線菌 │└────────────┘ ┃ │
│ ┃ └─────────────────┘ 原核生物:核膜がない ┃ │
│ ┃ 単細胞生物 真核生物:核膜がある ┃ │
│ ┗━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┛ │
│┌────────────────────────────────────────────────────┐│
││ 真核生物 ││
││┏━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┓││
││┃ 原生生物 ┃││
││┃┌────────────────────────────────────────────────┐┃││
││┃│ 植物性 │┃││
││┃│ ┌─────────────────────────┐ │┃││
││┃│ │ 藻類 │ │┃││
││┃│ │┌────────┐┌────┐┌───────┐│ │┃││
││┃│ ││ 緑藻類 ││紅藻類 ││ 褐藻類 ││ │┃││
││┃│ ││アオノリ、アオサ││テングサ││コンブ、ワカメ││ │┃││
││┃│ │└────────┘└────┘└───────┘│ │┃││
││┃│ └─────────────────────────┘ │┃││
││┃│┌──────┐┌──────┐┌──────┐┌────┐┌───────┐┌───────┐│┃││
││┃││ミドリムシ類││ 渦鞭毛藻類 ││ 珪藻類 ││ 卵菌類 ││ 変形菌類 ││ 細胞性粘菌類 ││┃││
││┃││ミドリムシ ││ ツノモ ││ハネケイソウ││ミズカビ││ムラサキホコリ││タマホコリカビ││┃││
││┃│└──────┘│ヤコウチュウ│└──────┘└────┘└───────┘└───────┘│┃││
││┃│ ↑ └──────┘ ↑ │┃││
││┃│ │ └真正粘菌 │┃││
││┃│ └鞭毛運動をする動物的性質を持ちながら、同時に植物として葉緑体を持ち光合成を行う「単細胞生物」│┃││
││┃│ │┃││
││┃│ クロロフィルa+b:ミドリムシ類、緑藻類、植物 │┃││
││┃│ クロロフィルa :紅藻類 │┃││
││┃│ クロロフィルa+c:渦鞭毛藻類、珪藻類、褐藻類 │┃││
││┃└────────────────────────────────────────────────┘┃││
││┃ ┌───────────────────────────────┐ ┃││
││┃ │ 動物性 │ ┃││
││┃ │┌────┐┌───────┐┌─────┐┌───────┐│ ┃││
││┃ ││肉質虫類││ 鞭毛虫類 ││ 繊毛虫類 ││ 胞子虫類 ││ ┃││
││┃ ││アメーバ││トリパノソーマ││ゾウリムシ││マラリア病原虫││ ┃││
││┃ │└────┘└───────┘└─────┘└───────┘│ ┃││
││┃ │ 単細胞生物 │ ┃││
││┃ └───────────────────────────────┘ ┃││
││┃ 藻類:酸素発生型光合成生物で地上生息以外の総称 ┃││
││┃ 水生・海生生息 ┃││
││┃ バクテリアに属する藻類(単細胞生物) ┃││
││┃ シアノバクテリア、藍藻、藍色細菌 ┃││
││┃ 真核生物に属する藻類(単細胞生物、多細胞生物) ┃││
││┃ ┃││
││┃ 菌類:ほとんどが多細胞生物、酵母は単細胞生物 ┃││
││┃ 地衣類は菌類と藻類の共生生物 ┃││
││┗━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┛││
││┏━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┓││
││┃ 植物 ┃││
││┃ ┌─────────────────────┐ ┃││
││┃ │ コケ植物 │ ┃││
││┃ │┌─────┐┌─────┐┌─────┐│ ┃││
││┃ ││ コケ類 ││ツノゴケ類││ 鮮類 ││ ┃││
││┃ ││ゼニゴケ ││ツノゴケ ││ミズゴケ ││ ┃││
││┃ │└─────┘└─────┘└─────┘│ ┃││
││┃ └─────────────────────┘ ┃││
││┃ ┌──────────────────────────────────────────────┐ ┃││
││┃ │ 維管束植物 │ ┃││
││┃ │ ┌──────────────────────────────┐ │ ┃││
││┃ │ │ シダ植物 │ │ ┃││
││┃ │ │┌─────┐┌────┐┌──────┐┌───────┐│ │ ┃││
││┃ │ ││ シダ類 ││トクサ類││マツバラン類││ヒゲノカズラ類││ │ ┃││
││┃ │ ││ゼンマイ ││トクサ ││マツバラン ││ヒゲノカズラ ││ │ ┃││
││┃ │ │└─────┘└────┘└──────┘└───────┘│ │ ┃││
││┃ │ └──────────────────────────────┘ │ ┃││
││┃ │┌────────────────────────────────────────────┐│ ┃││
││┃ ││ 種子植物 ││ ┃││
││┃ ││┌──────────────────────┐┌──────────────────┐││ ┃││
││┃ │││ 裸子植物 ││ 被子植物 │││ ┃││
││┃ │││┌──────┐┌─────┐┌─────┐││┌──────┐┌────────┐│││ ┃││
││┃ ││││ 球果類 ││ ソテツ類 ││イチョウ類││││ 単子葉類 ││ 双子葉類 ││││ ┃││
││┃ ││││スギ、ヒノキ││ ソテツ ││イチョウ ││││イネ、コムギ││サクラ、アサガオ││││ ┃││
││┃ │││└──────┘└─────┘└─────┘││└──────┘└────────┘│││ ┃││
││┃ ││└──────────────────────┘└──────────────────┘││ ┃││
││┃ │└────────────────────────────────────────────┘│ ┃││
││┃ │ 種子で増える:裸子植物・被子植物 │ ┃││
││┃ └──────────────────────────────────────────────┘ ┃││
││┃ 胞子で増える:藻類・菌類・コケ植物・シダ植物、維管束なし=コケ植物、維管束あり=シダ植物 ┃││
││┗━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┛││
││┏━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┓││
││┃ 菌類 ┃││
││┃ ┌──────┐┌─────┐┌──────┐┌──────┐┌───────┐ ┃││
││┃ │ 担子菌類 ││不完全菌類││ 子嚢菌類 ││ 接合菌類 ││ ツボカビ類 │ ┃││
││┃ │ シイタケ ││アオカビ ││アカパンカビ││クモノスカビ││カエルツボカビ│ ┃││
││┃ └──────┘└─────┘└──────┘└──────┘└───────┘ ┃││
││┃ ツボカビ類:鞭毛あり ┃││
││┃ 他 :鞭毛なし ┃││
││┃ カビの姿=菌糸+胞子 ┃││
││┃ キノコ=子実体+菌糸体+胞子 ┃││
││┗━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┛││
││┏━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┓││
││┃ 動物 ┃││
││┃ ┌────────────────┐ ┃││
││┃ │ 無胚葉 │ ┃││
││┃ │┌──────────────┐│ ┃││
││┃ ││ 海綿動物 ││ ┃││
││┃ ││カイロオウドウケツ、カイメン││ ┃││
││┃ │└──────────────┘│ ┃││
││┃ └────────────────┘ ┃││
││┃ ┌──────────────────────────────┐ ┃││
││┃ │ 二胚葉 │ ┃││
││┃ │ ┌─────┐┌───────────┐ │ ┃││
││┃ │ │ 有櫛動物 ││ 刺胞動物 │ │有櫛動物:刺胞なし┃││
││┃ │ │クシクラゲ││サンゴ、イソギンチャク│ │刺胞動物:刺胞あり┃││
││┃ │ └─────┘└───────────┘ │ ┃││
││┃ │ ┌───────────┐ │ ┃││
││┃ │ │ 平板動物 │ │ ┃││
││┃ │ │ センモウヒラムシ │ │ ┃││
││┃ │ └───────────┘ │ ┃││
││┃ └──────────────────────────────┘ ┃││
││┃ ┌──────────────────────────────────────────┐ ┃││
││┃ │ 三胚葉 │ ┃││
││┃ │┌────────────────────────────────────────┐│ ┃││
││┃ ││ 旧口動物 ││ ┃││
││┃ ││ ┌──────────────────────────────────┐ ││ ┃││
││┃ ││ │ 無体腔 │ ││ ┃││
││┃ ││ │┌────────────────────────────────┐│ ││ ┃││
││┃ ││ ││ 扁形動物 ││ ││ ┃││
││┃ ││ ││┌──────┐┌──────┐┌──────┐┌──────┐││ ││ ┃││
││┃ ││ │││ 渦虫類 ││ 吸虫類 ││ 単生類 ││ 条虫類 │││ ││ ┃││
││┃ ││ │││プラナリア ││ ジストマ ││フタゴムシ ││サナダムシ │││ ││ ┃││
││┃ ││ ││└──────┘└──────┘└──────┘└──────┘││ ││ ┃││
││┃ ││ │└────────────────────────────────┘│ ││ ┃││
││┃ ││ └──────────────────────────────────┘ ││ ┃││
││┃ ││ ┌────────────────────┐ ││ ┃││
││┃ ││ │ 偽体腔 │ ││ ┃││
││┃ ││ │┌────────┐┌────────┐│ ││ ┃││
││┃ ││ ││ 冠輪動物 ││ 脱皮動物 ││ ││ ┃││
││┃ ││ ││┌──────┐││┌──────┐││ ││ ┃││
││┃ ││ │││ 輪形動物 ││││ 線形動物 │││ ││ ┃││
││┃ ││ │││ツボワムシ ││││カイチュウ │││ ││ ┃││
││┃ ││ │││ネズミワムシ││││線虫、鞭虫 │││ ││ ┃││
││┃ ││ ││└──────┘││└──────┘││ ││ ┃││
││┃ ││ │└────────┘└────────┘│ ││ ┃││
││┃ ││ └────────────────────┘ ││ ┃││
││┃ ││┌──────────────────────────────────────┐││ ┃││
││┃ │││ 真体腔 │││ ┃││
││┃ │││┌────────────┐┌──────────────────────┐│││ ┃││
││┃ ││││ 冠輪動物 ││ 脱皮動物 ││││ ┃││
││┃ ││││ ┌──────┐ ││┌────────────────────┐││││ ┃││
││┃ ││││ │ 環形動物 │ │││ 節足動物 │││││ ┃││
││┃ ││││ │ヒル、ミミズ│ │││ ┌────────────┐ │││││ ┃││
││┃ ││││ └──────┘ │││ │ 鋏角類 │ │││││ ┃││
││┃ ││││┌──────────┐│││ │クモ、サソリ、カブトガニ│ │││││ ┃││
││┃ │││││ 軟体動物 ││││ └────────────┘ │││││ ┃││
││┃ │││││┌────────┐││││ ┌────────────┐ │││││ ┃││
││┃ ││││││ 二枚貝類 │││││ │ 多足類 │ │││││ ┃││
││┃ ││││││ハマグリ、アサリ│││││ │ ムカデ、ゲジ、ヤスデ │ │││││ ┃││
││┃ │││││└────────┘││││ └────────────┘ │││││ ┃││
││┃ │││││┌────────┐││││ ┌────────────────┐ │││││ ┃││
││┃ ││││││ 巻貝類 │││││ │ 甲殻類 │ │││││ ┃││
││┃ ││││││アワビ、サザエ │││││ │ エビ、カニ、ミジンコ、フジツボ │ │││││ ┃││
││┃ ││││││カタツムリ │││││ └────────────────┘ │││││ ┃││
││┃ │││││└────────┘││││┌──────────────────┐│││││ ┃││
││┃ │││││┌────────┐│││││ 六脚類 ││││││ ┃││
││┃ ││││││ 頭足類 ││││││┌────────────────┐││││││ ┃││
││┃ ││││││ タコ、イカ │││││││ 内顎類 │││││││ ┃││
││┃ ││││││ オウムガイ │││││││ トビムシ、コムシ、カマアシムシ │││││││ ┃││
││┃ │││││└────────┘│││││└────────────────┘││││││ ┃││
││┃ ││││└──────────┘││││ ┌──────────────┐ ││││││ ┃││
││┃ │││└────────────┘│││ │ 昆虫類 │ ││││││ ┃││
││┃ │││ │││ │カブトムシ、ゴミムシ │ ││││││ ┃││
││┃ │││ │││ │チョウ、ガ、トビケラ │ ││││││ ┃││
││┃ │││ │││ │ハエ、カ、アブ、ノミ、シラミ│ ││││││ ┃││
││┃ │││ │││ │スズメバチ、ミツバチ、アリ │ ││││││ ┃││
││┃ │││ │││ │セミ、カメムシ、アザミウマ │ ││││││ ┃││
││┃ │││ │││ │バッタ、キリギリス、コオロギ│ ││││││ ┃││
││┃ │││ │││ │カマキリ、カゲロウ、トンボ │ ││││││ ┃││
││┃ │││ │││ │ゴキブリ、シロアリ │ ││││││ ┃││
││┃ │││ │││ └──────────────┘ ││││││ ┃││
││┃ │││ ││└──────────────────┘│││││ ┃││
││┃ │││ │└────────────────────┘││││ ┃││
││┃ │││ └──────────────────────┘│││ ┃││
││┃ ││└──────────────────────────────────────┘││ ┃││
││┃ │└────────────────────────────────────────┘│ ┃││
││┃ │┌────────────────────────────────────────┐│ ┃││
││┃ ││ 新口動物 ││ ┃││
││┃ ││ ┌────┐┌───────────────┐ ││ ┃││
││┃ ││ │毛顎動物││ 棘皮動物 │ ││ ┃││
││┃ ││ │ヤムシ ││ウミユリ、ヒトデ、ウニ、ナマコ│ ││ ┃││
││┃ ││ └────┘└───────────────┘ ││ ┃││
││┃ ││┌──────────────────────────────────────┐││ ┃││
││┃ │││ 脊索動物 │││ ┃││
││┃ │││ ┌──────────────────┐ │││ ┃││
││┃ │││ │ 原索動物 │ │││ ┃││
││┃ │││ │┌───────┐┌───────┐│ │││ ┃││
││┃ │││ ││ 頭索類 ││ 尾索類 ││ │││ ┃││
││┃ │││ ││ ナメクジウオ ││ ホヤ ││ │││ ┃││
││┃ │││ │└───────┘└───────┘│ │││ ┃││
││┃ │││ └──────────────────┘ │││ ┃││
││┃ │││┌────────────────────────────────────┐│││ ┃││
││┃ ││││ 脊椎動物 ││││ ┃││
││┃ ││││ ┌──────┐ ││││ ┃││
││┃ ││││ │ 無顎類 │ ││││ ┃││
││┃ ││││ │ヤツメウナギ│ ││││ ┃││
││┃ ││││ └──────┘ ││││ ┃││
││┃ ││││┌──────────────────────────────────┐││││ ┃││
││┃ │││││ 顎口類 │││││ ┃││
││┃ │││││ ┌──────┐ │││││ ┃││
││┃ │││││ │ 軟骨魚類 │ │││││ ┃││
││┃ │││││ │サメ、エイ │ │││││ ┃││
││┃ │││││ └──────┘ │││││ ┃││
││┃ │││││┌────────────────────────────────┐│││││ ┃││
││┃ ││││││ 硬骨魚類 ││││││ ┃││
││┃ ││││││ ┌──────┐ ││││││ ┃││
││┃ ││││││ │ 条鰭類 │ ││││││ ┃││
││┃ ││││││ │コイ、タイ │ ││││││ ┃││
││┃ ││││││ └──────┘ ││││││ ┃││
││┃ ││││││┌──────────────────────────────┐││││││ ┃││
││┃ │││││││ 肉鰭類 │││││││ ┃││
││┃ │││││││ ┌───────┐┌─────┐ │││││││ ┃││
││┃ │││││││ │シーラカンス類││ 肺魚類 │ │││││││ ┃││
││┃ │││││││ │ラティメリア ││ハイギョ │ │││││││ ┃││
││┃ │││││││ └───────┘└─────┘ │││││││ ┃││
││┃ │││││││┌────────────────────────────┐│││││││ ┃││
││┃ ││││││││ 四足類 ││││││││ ┃││
││┃ ││││││││┌───┐┌─────────────────────┐││││││││ ┃││
││┃ │││││││││両生類││ 羊膜類 │││││││││ ┃││
││┃ │││││││││カエル││┌─────┐┌─────┐┌─────┐│││││││││ ┃││
││┃ │││││││││イモリ│││ 爬虫類 ││ 鳥類 ││ 哺乳類 ││││││││││ ┃││
││┃ ││││││││└───┘││ヘビ、ワニ││ハト、タカ││ヒト、イヌ││││││││││ ┃││
││┃ ││││││││ ││トカゲ ││スズメ ││クジラ ││││││││││ ┃││
││┃ ││││││││ │└─────┘└─────┘└─────┘│││││││││ ┃││
││┃ ││││││││ └─────────────────────┘││││││││ ┃││
││┃ │││││││└────────────────────────────┘│││││││ ┃││
││┃ ││││││└──────────────────────────────┘││││││ ┃││
││┃ │││││└────────────────────────────────┘│││││ ┃││
││┃ ││││└──────────────────────────────────┘││││ ┃││
││┃ │││└────────────────────────────────────┘│││ ┃││
││┃ ││└──────────────────────────────────────┘││ ┃││
││┃ │└────────────────────────────────────────┘│ ┃││
││┃ │ │ ┃││
││┃ │ 旧口動物・新口動物 │ ┃││
││┃ │ 体壁と消化管の隙間を体腔と呼び、体腔が内胚葉で包まれているものを真体腔と呼ぶ │ ┃││
││┃ │ 初期胚の体内を貫通する腸管が貫入した入口(先にできた口なので原口とか前口と呼ぶ)と │ ┃││
││┃ │ 初期胚の体内を貫通した腸管が反対側に出た出口(後にできた口なので後口と呼ぶ)が各々 │ ┃││
││┃ │ 何になるかによって旧口(前口)か新口(後口)かに分ける │ ┃││
││┃ │ │ ┃││
││┃ │ 入口が口になり、出口が肛門になるのが旧口(前口) … 入口:旧い口 │ ┃││
││┃ │ 出口が口になり、入口が肛門になるのが新口(後口) … 出口:新しい口 │ ┃││
││┃ │ │ ┃││
││┃ └──────────────────────────────────────────┘ ┃││
││┗━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━┛││
│└────────────────────────────────────────────────────┘│
└──────────────────────────────────────────────────────┘
・------------------------------------------------------------------------------------------------------------------------------・
:起源生物┬原核生物┬真正細菌(バクテリア) シアノバクテリア(藍藻、藍色細菌) :
: │ │ 紅色硫黄細菌、亜硫酸菌、枯草菌 :
: │ │ 大腸菌、乳酸菌、放線菌 :
: │ │----------------------------------------------------------------------------------------------------------:
: │ └古細菌(アーキア) メタン生成菌、超好熱菌 :
: │ : 高度酸性菌、高度好塩菌 :
: │--------------------------------------------------------------------------------------------------------------------:
: └真核生物┬原生生物┬植物性┬藻類┬緑藻類 アオノリ、アオサ :
: : │ │ │ │----------------------------------------------------------------------------------:
: : │ │ │ ├紅藻類 テングサ :
: : │ │ │ │----------------------------------------------------------------------------------:
: : │ │ │ └褐藻類 コンブ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ ├ミドリムシ類 ミドリムシ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ ├渦鞭毛藻類 ツノモ、ヤコウチュウ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ ├珪藻類 ハネケイソウ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ ├卵菌類 ミズカビ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ ├変形菌類(真正粘菌) ムラサキホコリ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ └細胞性粘菌類 タマホコリカビ :
: : │ │------------------------------------------------------------------------------------------------:
: : │ └動物性┬肉質虫類 アメーバ :
: : │ : │----------------------------------------------------------------------------------------:
: : │ : ├鞭毛虫類 トリパノソーマ :
: : │ : │----------------------------------------------------------------------------------------:
: : │ : ├繊毛虫類 ゾウリムシ :
: : │ : │----------------------------------------------------------------------------------------:
: : │ : └胞子虫類 マラリア病原虫 :
: : │----------------------------------------------------------------------------------------------------------:
: : ├植物┬コケ植物─┬コケ類 ゼニゴケ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ ├ツノゴケ類 ツノゴケ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ └鮮類 ミズゴケ :
: : │ │----------------------------------------------------------------------------------------------------:
: : │ └維管束植物┬シダ植物┬シダ類 ゼンマイ、ウラジロ、ワラビ :
: : │ : │ │------------------------------------------------------------------------------:
: : │ : │ ├トクサ類 トクサ :
: : │ : │ │------------------------------------------------------------------------------:
: : │ : │ ├マツバラン類 マツバラン :
: : │ : │ │------------------------------------------------------------------------------:
: : │ : │ └ヒゲノカズラ類 ヒゲノカズラ :
: : │ : │----------------------------------------------------------------------------------------:
: : │ : └種子植物┬裸子植物┬球果類 スギ、ヒノキ、メタセコイア :
: : │ : : │ │--------------------------------------------------------------------:
: : │ : : │ ├ソテツ類 ソテツ :
: : │ : : │ │--------------------------------------------------------------------:
: : │ : : │ └イチョウ類 イチョウ :
: : │ : : │------------------------------------------------------------------------------:
: : │ : : └被子植物┬双子葉植物┬合弁花類 アサガオ、ツツジ、タンポポ :
: : │ : : : │ │--------------------------------------------------------:
: : │ : : : │ └離弁花類 アブラナ、エンドウ、サクラ :
: : │ : : : │--------------------------------------------------------------------:
: : │ : : : └単子葉植物 イネ、コムギ、ショウガ、ラン :
: : ├----------------------------------------------------------------------------------------------------------:
: : ├菌類┬担子菌類 シイタケ、ヒトヨタケ :
: : │ │----------------------------------------------------------------------------------------------------:
: : │ ├不完全菌類 アオカビ :
: : │ │----------------------------------------------------------------------------------------------------:
: : │ ├子嚢菌類 アカパンカビ、酵母、冬虫夏草 :
: : │ │----------------------------------------------------------------------------------------------------:
: : │ ├接合菌類 クモノスカビ :
: : │ │----------------------------------------------------------------------------------------------------:
: : │ └ツボカビ類 カエルツボカビ :
: : │----------------------------------------------------------------------------------------------------------:
: : └動物┬無胚葉─海綿動物 カイロウドウケツ、カイメン :
: : : │----------------------------------------------------------------------------------------------------:
: : : ├二胚葉┬有櫛動物(刺胞なし) クシクラゲ :
: : : │ │--------------------------------------------------------------------------------------------:
: : : │ ├刺胞動物(刺胞あり) サンゴ、イソギンチャク :
: : : │ │--------------------------------------------------------------------------------------------:
: : : │ └平板動物 センモウヒラムシ :
: : : │----------------------------------------------------------------------------------------------------:
: : : └三胚葉┬旧口動物┬無体腔─扁形動物┬渦虫類 プラナリア :
: : : : │ │ │----------------------------------------------------------------:
: : : : │ │ ├吸虫類 ジストマ :
: : : : │ │ │----------------------------------------------------------------:
: : : : │ │ ├単生類 フタゴムシ :
: : : : │ │ │----------------------------------------------------------------:
: : : : │ │ └条虫類 サナダムシ :
: : : : │ │----------------------------------------------------------------------------------:
: : : : │ ├偽体腔┬冠輪動物─輪形動物 ツボワムシ、ネズミワムシ :
: : : : │ │ │--------------------------------------------------------------------------:
: : : : │ │ └脱皮動物─線形動物 カイチュウ、線虫、鞭虫 :
: : : : │ │----------------------------------------------------------------------------------:
: : : : │ └真体腔┬冠輪動物┬環形動物 ヒル、ミミズ :
: : : : │ : │ │----------------------------------------------------------------:
: : : : │ : │ └軟体動物┬二枚貝類 ハマグリ、アサリ :
: : : : │ : │ : │------------------------------------------------------:
: : : : │ : │ : ├巻貝類 アワビ、サザエ、カタツムリ :
: : : : │ : │ : │------------------------------------------------------:
: : : : │ : │ : └頭足類 タコ、イカ、オウムガイ :
: : : : │ : │--------------------------------------------------------------------------:
: : : : │ : └脱皮動物─節足動物┬鋏角類 クモ、サソリ、カブトガニ :
: : : : │ : : │------------------------------------------------------:
: : : : │ : : ├多足類 ムカデ、ゲジ、ヤスデ :
: : : : │ : : │------------------------------------------------------:
: : : : │ : : ├甲殻類 エビ、カニ、ミジンコ、フジツボ :
: : : : │ : : │------------------------------------------------------:
: : : : │ : : └六脚類┬内顎類 トビムシ、コムシ、カマアシムシ :
: : : : │ : : : │----------------------------------------------:
: : : : │ : : : └昆虫類 カブトムシ、ゴミムシ :
: : : : │ : : : : チョウ、ガ、トビケラ :
: : : : │ : : : : ハエ、カ、アブ、ノミ、シラミ :
: : : : │ : : : : スズメバチ、ミツバチ、アリ :
: : : : │ : : : : セミ、カメムシ、アザミウマ :
: : : : │ : : : : バッタ、キリギリス、コオロギ :
: : : : │ : : : : カマキリ、カゲロウ、トンボ :
: : : : │ : : : : ゴキブリ、シロアリ :
: : : : │--------------------------------------------------------------------------------------------:
: : : : └新口動物┬毛顎動物 ヤムシ :
: : : : : │----------------------------------------------------------------------------------:
: : : : : ├棘皮動物 ウミユリ、ヒトデ、ウニ、ナマコ :
: : : : : │----------------------------------------------------------------------------------:
: : : : : └脊索動物┬原索動物┬頭索類 ナメクジウオ :
: : : : : : : │--------------------------------------------------------------:
: : : : : : │ └尾索類 ホヤ :
: : : : : : │------------------------------------------------------------------------:
: : : : : : └脊椎動物┬無顎類 ヤツメウナギ :
: : : : : : : │--------------------------------------------------------------:
: : : : : : : └顎口類┬軟骨魚類 サメ、エイ :
: : : : : : : : │------------------------------------------------------:
: : : : : : : : └硬骨魚類┬条鰭類 コイ、タイ :
: : : : : : : : : │--------------------------------------------:
: : : : : : : : : └肉鰭類┬シーラカンス類 ラティメリア :
: : : : : : : : : : │------------------------------------:
: : : : : : : : : : ├肺魚類 ハイギョ :
: : : : : : : : : : │------------------------------------:
: : : : : : : : : : └四足類┬両生類 カエル、イモリ :
: : : : : : : : : : : │----------------------------:
: : : : : : : : : : : └羊膜類┬爬虫類 ヘビ、ワニ :
: : : : : : : : : : : : │--------------------:
: : : : : : : : : : : : ├鳥類 ハト、タカ :
: : : : : : : : : : : : │--------------------:
: : : : : : : : : : : : └哺乳類 ヒト、イヌ :
・------------------------------------------------------------------------------------------------------------------------------・
:起源生物┬原核生物┬真正細菌(バクテリア) シアノバクテリア(藍藻、藍色細菌) :
: │ │ 紅色硫黄細菌、亜硫酸菌、枯草菌 :
: │ │ 大腸菌、乳酸菌、放線菌 :
: │ │----------------------------------------------------------------------------------------------------------:
: │ └古細菌(アーキア) メタン生成菌、超好熱菌 :
: │ : 高度酸性菌、高度好塩菌 :
: │--------------------------------------------------------------------------------------------------------------------:
: └真核生物┬原生生物┬植物性┬藻類┬緑藻類 アオノリ、アオサ :
: : │ │ │ │----------------------------------------------------------------------------------:
: : │ │ │ ├紅藻類 テングサ :
: : │ │ │ │----------------------------------------------------------------------------------:
: : │ │ │ └褐藻類 コンブ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ ├ミドリムシ類 ミドリムシ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ ├渦鞭毛藻類 ツノモ、ヤコウチュウ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ ├珪藻類 ハネケイソウ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ ├卵菌類 ミズカビ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ ├変形菌類(真正粘菌) ムラサキホコリ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ └細胞性粘菌類 タマホコリカビ :
: : │ │------------------------------------------------------------------------------------------------:
: : │ └動物性┬肉質虫類 アメーバ :
: : │ : │----------------------------------------------------------------------------------------:
: : │ : ├鞭毛虫類 トリパノソーマ :
: : │ : │----------------------------------------------------------------------------------------:
: : │ : ├繊毛虫類 ゾウリムシ :
: : │ : │----------------------------------------------------------------------------------------:
: : │ : └胞子虫類 マラリア病原虫 :
: : │----------------------------------------------------------------------------------------------------------:
: : ├植物┬コケ植物─┬コケ類 ゼニゴケ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ ├ツノゴケ類 ツノゴケ :
: : │ │ │----------------------------------------------------------------------------------------:
: : │ │ └鮮類 ミズゴケ :
: : │ │----------------------------------------------------------------------------------------------------:
: : │ └維管束植物┬シダ植物┬シダ類 ゼンマイ、ウラジロ、ワラビ :
: : │ : │ │------------------------------------------------------------------------------:
: : │ : │ ├トクサ類 トクサ :
: : │ : │ │------------------------------------------------------------------------------:
: : │ : │ ├マツバラン類 マツバラン :
: : │ : │ │------------------------------------------------------------------------------:
: : │ : │ └ヒゲノカズラ類 ヒゲノカズラ :
: : │ : │----------------------------------------------------------------------------------------:
: : │ : └種子植物┬裸子植物┬球果類 スギ、ヒノキ、メタセコイア :
: : │ : : │ │--------------------------------------------------------------------:
: : │ : : │ ├ソテツ類 ソテツ :
: : │ : : │ │--------------------------------------------------------------------:
: : │ : : │ └イチョウ類 イチョウ :
: : │ : : │------------------------------------------------------------------------------:
: : │ : : └被子植物┬双子葉植物┬合弁花類 アサガオ、ツツジ、タンポポ :
: : │ : : : │ │--------------------------------------------------------:
: : │ : : : │ └離弁花類 アブラナ、エンドウ、サクラ :
: : │ : : : │--------------------------------------------------------------------:
: : │ : : : └単子葉植物 イネ、コムギ、ショウガ、ラン :
: : ├----------------------------------------------------------------------------------------------------------:
: : ├菌類┬担子菌類 シイタケ、ヒトヨタケ :
: : │ │----------------------------------------------------------------------------------------------------:
: : │ ├不完全菌類 アオカビ :
: : │ │----------------------------------------------------------------------------------------------------:
: : │ ├子嚢菌類 アカパンカビ、酵母、冬虫夏草 :
: : │ │----------------------------------------------------------------------------------------------------:
: : │ ├接合菌類 クモノスカビ :
: : │ │----------------------------------------------------------------------------------------------------:
: : │ └ツボカビ類 カエルツボカビ :
: : │----------------------------------------------------------------------------------------------------------:
: : └動物┬無胚葉─海綿動物 カイロウドウケツ、カイメン :
: : : │----------------------------------------------------------------------------------------------------:
: : : ├二胚葉┬有櫛動物(刺胞なし) クシクラゲ :
: : : │ │--------------------------------------------------------------------------------------------:
: : : │ ├刺胞動物(刺胞あり) サンゴ、イソギンチャク :
: : : │ │--------------------------------------------------------------------------------------------:
: : : │ └平板動物 センモウヒラムシ :
: : : │----------------------------------------------------------------------------------------------------:
: : : └三胚葉┬旧口動物┬無体腔─扁形動物┬渦虫類 プラナリア :
: : : : │ │ │----------------------------------------------------------------:
: : : : │ │ ├吸虫類 ジストマ :
: : : : │ │ │----------------------------------------------------------------:
: : : : │ │ ├単生類 フタゴムシ :
: : : : │ │ │----------------------------------------------------------------:
: : : : │ │ └条虫類 サナダムシ :
: : : : │ │----------------------------------------------------------------------------------:
: : : : │ ├偽体腔┬冠輪動物─輪形動物 ツボワムシ、ネズミワムシ :
: : : : │ │ │--------------------------------------------------------------------------:
: : : : │ │ └脱皮動物─線形動物 カイチュウ、線虫、鞭虫 :
: : : : │ │----------------------------------------------------------------------------------:
: : : : │ └真体腔┬冠輪動物┬環形動物 ヒル、ミミズ :
: : : : │ : │ │----------------------------------------------------------------:
: : : : │ : │ └軟体動物┬二枚貝類 ハマグリ、アサリ :
: : : : │ : │ : │------------------------------------------------------:
: : : : │ : │ : ├巻貝類 アワビ、サザエ、カタツムリ :
: : : : │ : │ : │------------------------------------------------------:
: : : : │ : │ : └頭足類 タコ、イカ、オウムガイ :
: : : : │ : │--------------------------------------------------------------------------:
: : : : │ : └脱皮動物─節足動物┬鋏角類 クモ、サソリ、カブトガニ :
: : : : │ : : │------------------------------------------------------:
: : : : │ : : ├多足類 ムカデ、ゲジ、ヤスデ :
: : : : │ : : │------------------------------------------------------:
: : : : │ : : ├甲殻類 エビ、カニ、ミジンコ、フジツボ :
: : : : │ : : │------------------------------------------------------:
: : : : │ : : └六脚類┬内顎類 トビムシ、コムシ、カマアシムシ :
: : : : │ : : : │----------------------------------------------:
: : : : │ : : : └昆虫類 カブトムシ、ゴミムシ :
: : : : │ : : : : チョウ、ガ、トビケラ :
: : : : │ : : : : ハエ、カ、アブ、ノミ、シラミ :
: : : : │ : : : : スズメバチ、ミツバチ、アリ :
: : : : │ : : : : セミ、カメムシ、アザミウマ :
: : : : │ : : : : バッタ、キリギリス、コオロギ :
: : : : │ : : : : カマキリ、カゲロウ、トンボ :
: : : : │ : : : : ゴキブリ、シロアリ :
: : : : │--------------------------------------------------------------------------------------------:
: : : : └新口動物┬毛顎動物 ヤムシ :
: : : : : │----------------------------------------------------------------------------------:
: : : : : ├棘皮動物 ウミユリ、ヒトデ、ウニ、ナマコ :
: : : : : │----------------------------------------------------------------------------------:
: : : : : └脊索動物┬原索動物┬頭索類 ナメクジウオ :
: : : : : : : │--------------------------------------------------------------:
: : : : : : │ └尾索類 ホヤ :
: : : : : : │------------------------------------------------------------------------:
: : : : : : └脊椎動物┬無顎類 ヤツメウナギ :
: : : : : : : │--------------------------------------------------------------:
: : : : : : : └顎口類┬軟骨魚類 サメ、エイ :
: : : : : : : : │------------------------------------------------------:
: : : : : : : : └硬骨魚類┬条鰭類 コイ、タイ :
: : : : : : : : : │--------------------------------------------:
: : : : : : : : : └肉鰭類┬シーラカンス類 ラティメリア :
: : : : : : : : : : │------------------------------------:
: : : : : : : : : : ├肺魚類 ハイギョ :
: : : : : : : : : : │------------------------------------:
: : : : : : : : : : └四足類┬両生類 カエル、イモリ :
: : : : : : : : : : : │----------------------------:
: : : : : : : : : : : └羊膜類┬爬虫類 ヘビ、ワニ :
: : : : : : : : : : : : │--------------------:
: : : : : : : : : : : : ├鳥類 ハト、タカ :
: : : : : : : : : : : : │--------------------:
: : : : : : : : : : : : └哺乳類 ヒト、イヌ :
・------------------------------------------------------------------------------------------------------------------------------・
(参考)二界説 … 動物界:動いてえさをとるもの(動物性原生生物、動物)
植物界:動物ではないもの(原核生物、植物性原生生物、菌類、植物)
植物界:動物ではないもの(原核生物、植物性原生生物、菌類、植物)
| 生産者 | 生産者 | 植物:無機物から有機物を合成 |
| 消費者 | 消費者 | 動物:植物を食べて有機物を排泄 |
| 分解者 | 菌類:植物と栄養分をやりとりして共生 | |
| きのこ | 有機物を分解・吸収して無機物に戻す役割 |
生物全体:184万種 … 世界で確認されている生物種数
動物:130万種┬節足動物:109万種、全動物種の85%以上を占める、大部分が昆虫:100万種
植物: 37万種├軟体動物: 6万種
菌類: 14万種├脊索動物: 7万種┬魚類:3万種
藻類: 2万種└ その他: 8万種├鳥類:1万種
その他: 1万種 └両生類+爬虫類+哺乳類+その他:3万種
動物:130万種┬節足動物:109万種、全動物種の85%以上を占める、大部分が昆虫:100万種
植物: 37万種├軟体動物: 6万種
菌類: 14万種├脊索動物: 7万種┬魚類:3万種
藻類: 2万種└ その他: 8万種├鳥類:1万種
その他: 1万種 └両生類+爬虫類+哺乳類+その他:3万種
地球上の生物種数は推定約870万とする研究報告発表(2011) … 未知の生物を含む
陸上種650万、海洋種220万 → 陸上種の86%、海洋種の91%が未知種
国際自然保護連合はレッドリストを公開(2019) … 絶滅危惧種の数は28,338種
1975年~2000年の25年間に年平均4万種、13分間に1種の生き物が絶滅
環境省レッドリスト(日本の生物)
野生動物の3割が絶滅危惧種
野生植物の2割が絶滅危惧種
| 物質粒子 | 力の伝達粒子 | 力の作用先 | 力の種類 | 力の強弱:目安 | 力の作用範囲 |
|---|---|---|---|---|---|
| 原子 | 光子 | 原子核・電子 | 電磁気力 | 弱い:0.01 | 原子核からの距離の二乗に反比例 |
| 原子核 | 中間子 | 陽子・中性子 | 強い力(核力) | 強い:1 | 10-13cm:水素原子核(陽子サイズ) |
| 素粒子 | グルーオン | クォーク | 強い力(色の力) | ||
| 弱ボソン | フェルミ粒子 | 弱い力 | はるかに弱い:10-14~10-7 | 10-16cm:水素原子核の1/1000 |
物質粒子 :物の成り立ちの基本の粒子
力の伝達粒子:物を動かす力の基本の粒子 … 力は素粒子によって媒介される(交換:放出⇔吸収)
力の作用先 :複合粒子(陽子,中性子)、クォーク(u,d,s)、フェルミ粒子(クォーク,レプトン)
電磁気力 :原子同士を結び付け分子を作る力も電磁気力です
弱ボソン :弱い力はフェルミ粒子以外に、ハドロン(バリオン・メソン)にも作用する
力の伝達粒子:物を動かす力の基本の粒子 … 力は素粒子によって媒介される(交換:放出⇔吸収)
力の作用先 :複合粒子(陽子,中性子)、クォーク(u,d,s)、フェルミ粒子(クォーク,レプトン)
電磁気力 :原子同士を結び付け分子を作る力も電磁気力です
弱ボソン :弱い力はフェルミ粒子以外に、ハドロン(バリオン・メソン)にも作用する
核力:中間子はグルーオンが媒介する強い力の「強い残留力」を近接核子に伝える(近づき過ぎると斥力になる!)
核力は遠距離では引力ですが、至近距離では強い斥力になることが分かってきましたが、これを核力の斥力芯と呼びます
この斥力芯のおかげで、陽子や中性子が近づきすぎず、原子核は比較的低密度に保たれています(引力で潰れずに済む)
強い相互作用は、2つの範囲で観測でき、2つのフォースキャリア(中間子、グルーオン)により媒介される
大きいスケール(約1-3 fm)では、陽子と中性子(核子)を結合させて原子核を形成する力(中間子が運ぶ力)である
小さいスケール(核子の半径約0.8 fm以下)では、クォークを結合して陽子、中性子などのハドロン粒子を形成する力
である(グルーオンが運ぶ色の力(color force))
大きいスケールでは、中間子が陽子や中性子間を行き来するとき、
陽子や中性子の中のグルーオンがクォーク間を行き来して媒介される強い相互作用が核力を生みます
π中間子はクォーク○と反クォーク●の対で、グルーオンにより結びついている
大きいスケールでは、コイル状のバネのように強い力が働く:離れるほど引力が強くなり、近づくほど斥力が強くなる
色の力:色力線の閉じ込めで核子の内部でしか働かない、電磁気力:原子の大きさの数倍以内の範囲で働く
弱い力:粒子の種類を変える力など、力には以上3つの力のほか、マクロの世界を支配する「重力」がある
核力は遠距離では引力ですが、至近距離では強い斥力になることが分かってきましたが、これを核力の斥力芯と呼びます
この斥力芯のおかげで、陽子や中性子が近づきすぎず、原子核は比較的低密度に保たれています(引力で潰れずに済む)
|←─-陽子-─→| |←─中間子─→| |←─中性子─→|
|○⇔ℓℓℓℓℓℓ⇔○|⇔⇔⇔|○⇔ℓℓℓℓℓℓ⇔●|⇔⇔⇔|○⇔ℓℓℓℓℓℓ⇔○|
| グルーオン | | グルーオン | | グルーオン |
陽子・中性子の中の3つ目のクォーク○は省略|○⇔ℓℓℓℓℓℓ⇔○|⇔⇔⇔|○⇔ℓℓℓℓℓℓ⇔●|⇔⇔⇔|○⇔ℓℓℓℓℓℓ⇔○|
| グルーオン | | グルーオン | | グルーオン |
大きいスケール(約1-3 fm)では、陽子と中性子(核子)を結合させて原子核を形成する力(中間子が運ぶ力)である
小さいスケール(核子の半径約0.8 fm以下)では、クォークを結合して陽子、中性子などのハドロン粒子を形成する力
である(グルーオンが運ぶ色の力(color force))
大きいスケールでは、中間子が陽子や中性子間を行き来するとき、
陽子や中性子の中のグルーオンがクォーク間を行き来して媒介される強い相互作用が核力を生みます
π中間子はクォーク○と反クォーク●の対で、グルーオンにより結びついている
大きいスケールでは、コイル状のバネのように強い力が働く:離れるほど引力が強くなり、近づくほど斥力が強くなる
色の力:色力線の閉じ込めで核子の内部でしか働かない、電磁気力:原子の大きさの数倍以内の範囲で働く
弱い力:粒子の種類を変える力など、力には以上3つの力のほか、マクロの世界を支配する「重力」がある
[陽子のパートン構造]:陽子をより短い時間の尺度で観測すると、内部構造が現れる
強い相互作用によって、グルーオンの放射やクォーク・反クォークの対生成が起こり、
クォーク・反クォーク・グルーオンという素粒子(=パートン)の複合体になっている
強い力の基本結合:グルーオン放射、グルーオン分裂、グルーオン自己結合
グルーオンは自らが色荷をもっているため、グルーオン同士も相互作用します
強い相互作用によって、グルーオンの放射やクォーク・反クォークの対生成が起こり、
クォーク・反クォーク・グルーオンという素粒子(=パートン)の複合体になっている
強い力の基本結合:グルーオン放射、グルーオン分裂、グルーオン自己結合
グルーオンは自らが色荷をもっているため、グルーオン同士も相互作用します
| 相互作用 | クォーク | レプトン | 備考 | |
|---|---|---|---|---|
| 荷電レプトン | ニュートリノ | |||
| 電磁気力 | ○ | ○ | × | 荷電レプトン:(e±,μ±,τ±) |
| 強い力 | ○ | × | × | クォーク :(u,c,t),(d,s,b) |
| 弱い力 | ○ | ○ | ○ | ニュートリノ:(νe,νμ,ντ) |
| 重力 | ○ | ○ | ○ | ○:力を及ぼす、×:力を及ばさない |
レプトン:色荷無し、ニュートリノ:電荷無し
原子核と電子は、電気力を運ぶ光子をやりとり(放出⇔吸収)して引き合う
光子が互いに電気力を伝え、+の原子核と-の電子を電気力で引寄せさせる
マクロの世界 ミクロの世界
従来の電磁気学─────→場の量子化────→量子電気力学
電荷と電場の相互作用 力も飛び飛び 光子の交換
光子が互いに電気力を伝え、+の原子核と-の電子を電気力で引寄せさせる
マクロの世界 ミクロの世界
従来の電磁気学─────→場の量子化────→量子電気力学
電荷と電場の相互作用 力も飛び飛び 光子の交換
粒子同士は粒子間で力の伝達粒子をキャッチボールして力を伝えると考えます
キャッチボールを続けるには、ボールが届く相手のそばにいなければなりません
キャッチボールすること自体が粒子同士をつなぐ接着剤のような役目を果たします
力の伝達粒子がボールであれば、互いに向き合って受け取ると斥力になります
力の伝達粒子がブーメランであれば、背中合わせで受け取ると引力になります
力が引力になるか斥力になるかは、交換する仮想量子のスピンによって決まります
スピン1の光子は同符号の荷電粒子同士を反発させ異符号の荷電粒子同士を引寄せる
光子=ボール(斥力):++/--、光子=ブーメラン(引力):+-/-+
同符号の荷電粒子同士はエネルギーが高くなるので斥力が働く
異符号の荷電粒子同士はエネルギーが低くなるので引力が働く
物理現象はエネルギーが低い方向へと流れるため光子の交換
でエネルギーがより低い安定した状態に移行しようとします
光子の交換=量子化された場を介して相互作用するという意味(場が力を伝えている)
磁石の場合(光子は電磁気力を伝える)
電気の力+磁気の力=電磁気力
電流を導線に流すと導線のまわりに磁場ができる
磁石をコイルに出し入れするとコイルに電流が発生する(電磁誘導)
磁石は電子のスピン:電子が自転する(環状の電流)と磁石になる
電子1個にもN極とS極があり磁気を帯びています
磁石の場合、光子がN極の電子とS極の電子を行き来することで引力が生じます
磁石の場合、光子が同じ極の電子の間を行き来することで反発力(斥力)が生じます
キャッチボールを続けるには、ボールが届く相手のそばにいなければなりません
キャッチボールすること自体が粒子同士をつなぐ接着剤のような役目を果たします
力の伝達粒子がボールであれば、互いに向き合って受け取ると斥力になります
力の伝達粒子がブーメランであれば、背中合わせで受け取ると引力になります
力が引力になるか斥力になるかは、交換する仮想量子のスピンによって決まります
スピン1の光子は同符号の荷電粒子同士を反発させ異符号の荷電粒子同士を引寄せる
光子=ボール(斥力):++/--、光子=ブーメラン(引力):+-/-+
同符号の荷電粒子同士はエネルギーが高くなるので斥力が働く
異符号の荷電粒子同士はエネルギーが低くなるので引力が働く
物理現象はエネルギーが低い方向へと流れるため光子の交換
でエネルギーがより低い安定した状態に移行しようとします
光子の交換=量子化された場を介して相互作用するという意味(場が力を伝えている)
磁石の場合(光子は電磁気力を伝える)
電気の力+磁気の力=電磁気力
電流を導線に流すと導線のまわりに磁場ができる
磁石をコイルに出し入れするとコイルに電流が発生する(電磁誘導)
磁石は電子のスピン:電子が自転する(環状の電流)と磁石になる
電子1個にもN極とS極があり磁気を帯びています
磁石の場合、光子がN極の電子とS極の電子を行き来することで引力が生じます
磁石の場合、光子が同じ極の電子の間を行き来することで反発力(斥力)が生じます
q1q2 m1m2
F=k─── F=G───
r2 r2
k:クーロン定数 G:万有引力定数F=k─── F=G───
r2 r2
F:力、q:(±)電荷量、m:質量、r:距離
クーロンの法則の式は万有引力の法則の式に似ているが、万有引力には斥力がない
原子核の周りを回る電子の波は1周の軌道の長さが波長で割り切れる周回位相一致
原子内の電子の軌道 :K殻・L殻・M殻…各軌道グループ
原子核内の核子の軌道:Ⅰ殻・Ⅱ殻・Ⅲ殻…各軌道グループ
原子核の外側を回る複数電子も原子核の内側を回る複数核子も各軌道を回っている
各軌道には粒子の定員がありエネルギーの低い内側の軌道から順に満席にしていく
各電子殻に存在する軌道は、外側になるほど数が増え、K殻にはs軌道、L殻にはs,
p軌道、M殻にはs,p,d軌道、N殻にはs,p,d,f軌道 … が存在する
ジャンプ先の軌道はすぐ外側とは限らないが粒子で満席でなく空席がある条件では
特定軌道の粒子に光子を衝突させると、粒子は光子を吸収して、外側の軌道に移る
外側の軌道にジャンプした粒子は、暫くすると光子を放出して、内側の軌道に戻る
粒子が不連続に原子核の周回軌道を変える現象(ジャンプ)を量子飛躍と言います
原子内の電子の軌道や運動を調べる方法:原子分光法
原子核内の核子の軌道運動や自転運動、核子の集団運動を調べる方法:原子核分光法
・電子は放電や光のエネルギーを受けると、原子核から遠い外側の軌道に移る
・外側の軌道に移った電子が再び内側の軌道に移るとき、
内外の軌道のエネルギーの差を光子として放出する
外側の軌道→K殻…紫外線
外側の軌道→L殻…可視光線
外側の軌道→M殻…赤外線
高エネルギー粒子がK殻の電子に衝突すると、電子は原子の外に飛び出して、
K殻の軌道は空席ができ、外側のL殻やM殻の軌道の電子がK殻の軌道の
空席にジャンプし、それぞれの軌道のエネルギー差の分の光子を放出する
… KX線と言う特性X線
特定の光子(E=hν)を原子内に照射すると、外側軌道の電子が内側軌道
に移り、入射光子と同位相・同波長の放射光(E=hν)を放出する(誘導放出)
誘導放出を利用して位相と波長のそろった強い光、レーザー光が得られる
原子内の電子の軌道 :K殻・L殻・M殻…各軌道グループ
原子核内の核子の軌道:Ⅰ殻・Ⅱ殻・Ⅲ殻…各軌道グループ
原子核の外側を回る複数電子も原子核の内側を回る複数核子も各軌道を回っている
各軌道には粒子の定員がありエネルギーの低い内側の軌道から順に満席にしていく
各電子殻に存在する軌道は、外側になるほど数が増え、K殻にはs軌道、L殻にはs,
p軌道、M殻にはs,p,d軌道、N殻にはs,p,d,f軌道 … が存在する
ジャンプ先の軌道はすぐ外側とは限らないが粒子で満席でなく空席がある条件では
特定軌道の粒子に光子を衝突させると、粒子は光子を吸収して、外側の軌道に移る
外側の軌道にジャンプした粒子は、暫くすると光子を放出して、内側の軌道に戻る
粒子が不連続に原子核の周回軌道を変える現象(ジャンプ)を量子飛躍と言います
原子内の電子の軌道や運動を調べる方法:原子分光法
原子核内の核子の軌道運動や自転運動、核子の集団運動を調べる方法:原子核分光法
・電子は放電や光のエネルギーを受けると、原子核から遠い外側の軌道に移る
・外側の軌道に移った電子が再び内側の軌道に移るとき、
内外の軌道のエネルギーの差を光子として放出する
外側の軌道→K殻…紫外線
外側の軌道→L殻…可視光線
外側の軌道→M殻…赤外線
高エネルギー粒子がK殻の電子に衝突すると、電子は原子の外に飛び出して、
K殻の軌道は空席ができ、外側のL殻やM殻の軌道の電子がK殻の軌道の
空席にジャンプし、それぞれの軌道のエネルギー差の分の光子を放出する
… KX線と言う特性X線
特定の光子(E=hν)を原子内に照射すると、外側軌道の電子が内側軌道
に移り、入射光子と同位相・同波長の放射光(E=hν)を放出する(誘導放出)
誘導放出を利用して位相と波長のそろった強い光、レーザー光が得られる
電子のエネルギーは外側の軌道の電子ほど軌道特有のエネルギーが高い
原子核と電子の結合エネルギーは内側ほどエネルギーが高いので、回転
遠心力と平衡を保つために、近いほど速く回り、遠いほどゆっくり回る
原子番号が増えるほど電子を引きつける原子核の電気の力も大きくなる
化学反応では原子と原子が外側の電子を交換したり共有して分子を作る
原子核と電子の結合エネルギーは内側ほどエネルギーが高いので、回転
遠心力と平衡を保つために、近いほど速く回り、遠いほどゆっくり回る
原子番号が増えるほど電子を引きつける原子核の電気の力も大きくなる
化学反応では原子と原子が外側の電子を交換したり共有して分子を作る
TOP|
強い力(核力)
陽子と中性子は相互にπ±中間子を放出・吸収することによって核力が発生する
電荷の衣のやりとりで「陽子→中性子」「中性子→陽子」のように状態が変化する
→π+→
陽子 ⇔ 中性子
←π-←
陽子→π+→中性子 中性子→π-→陽子
↓ ↓ ↓ ↓
中性子 陽子 陽子 中性子
陽子同士や中性子同士はπ0中間子を放出・吸収することによって核力が発生する
陽子→π0→陽子 中性子→π0→中性子
原子核の中ではこれが繰り返されている(電荷の状態が変わっても核子の本質は不変)
陽子とマイナスの中間子がセットで中性子、中性子とプラスの中間子のセットで陽子
元:中間子が+の電荷を運んで陽子が中性子に-の電荷を運んで中性子が陽子になる
先:中間子が+の電荷を運んで中性子が陽子に-の電荷を運んで陽子が中性子になる
クォーク3個の質量は陽子1個の質量の1%以下
陽子の質量の99%は強い力のエネルギーによる質量←E=mc2で換算すると相当する
電荷の衣のやりとりで「陽子→中性子」「中性子→陽子」のように状態が変化する
→π+→
陽子 ⇔ 中性子
←π-←
陽子→π+→中性子 中性子→π-→陽子
↓ ↓ ↓ ↓
中性子 陽子 陽子 中性子
陽子同士や中性子同士はπ0中間子を放出・吸収することによって核力が発生する
陽子→π0→陽子 中性子→π0→中性子
原子核の中ではこれが繰り返されている(電荷の状態が変わっても核子の本質は不変)
陽子とマイナスの中間子がセットで中性子、中性子とプラスの中間子のセットで陽子
元:中間子が+の電荷を運んで陽子が中性子に-の電荷を運んで中性子が陽子になる
先:中間子が+の電荷を運んで中性子が陽子に-の電荷を運んで陽子が中性子になる
クォーク3個の質量は陽子1個の質量の1%以下
陽子の質量の99%は強い力のエネルギーによる質量←E=mc2で換算すると相当する
原子核の中では核子同士が中間子を交換して核力が生じていますが、核子が単体で存在
している場合でも、核子が中間子を放出して自己吸収するという過程が起こり得、この
過程では中間子が核子を取り巻く雲のように分布すると考えられる(中間子雲モデル)
陽子は一時的に中間子を放出し中性子に姿を変え、その後中間子を吸収し再び陽子に
戻ることができます
中間子雲モデルでは、中間子が雲のように中性子を覆う状態として陽子を捉えます
している場合でも、核子が中間子を放出して自己吸収するという過程が起こり得、この
過程では中間子が核子を取り巻く雲のように分布すると考えられる(中間子雲モデル)
陽子は一時的に中間子を放出し中性子に姿を変え、その後中間子を吸収し再び陽子に
戻ることができます
中間子雲モデルでは、中間子が雲のように中性子を覆う状態として陽子を捉えます
[パウリの排他律]:物質を構成する粒子は、同じ種類が1つの場所に同時には存在しえない
陽子(uud:uクォーク2個とdクォーク1個)、中性子(udd:uクォーク1個とdクォーク2個)
では、同じ種類のクォークが同じ場所で同時に重なり合って1つのハドロンを構成している
排他律違反を回避する色という属性が互いに異なる3色(赤、青、緑)のクォークを考えた
3種のクォークで構成する陽子や中性子などのバリオンは3色がそろえば白(無色)になる
補色であるシアン(反赤)、イエロー(反青)、マゼンタ(反緑)を反クォークの色として
メソンもやはり白と表現できる:赤クォークは反赤クオークと合体して、白いメソンになる
これでクォークに色があっても、バリオンやメソンには色がないことが自然に表現できます
陽子(uud:uクォーク2個とdクォーク1個)、中性子(udd:uクォーク1個とdクォーク2個)
では、同じ種類のクォークが同じ場所で同時に重なり合って1つのハドロンを構成している
排他律違反を回避する色という属性が互いに異なる3色(赤、青、緑)のクォークを考えた
3種のクォークで構成する陽子や中性子などのバリオンは3色がそろえば白(無色)になる
補色であるシアン(反赤)、イエロー(反青)、マゼンタ(反緑)を反クォークの色として
メソンもやはり白と表現できる:赤クォークは反赤クオークと合体して、白いメソンになる
これでクォークに色があっても、バリオンやメソンには色がないことが自然に表現できます
|
陽子(uud)の色の例:u赤u青d緑 中性子(udd)の色の例:u赤d青d緑 π+中間子(u反d)の色の例:u赤d反赤 |  |
クオーク同士を結び付けているのは色の力で、色の力を伝えるのはグルーオン
3色(赤、青、緑)のクォーク→グルーオン→3色(緑、青、赤)のクォーク
中間子(赤、反赤)のクォーク→グルーオン→中間子(青、反青)のクォーク
クオークは色の組(受取る色と渡す色)をもつグルーオンを通して色を交換する
3色(赤、青、緑)のクォーク→グルーオン→3色(緑、青、赤)のクォーク
中間子(赤、反赤)のクォーク→グルーオン→中間子(青、反青)のクォーク
クオークは色の組(受取る色と渡す色)をもつグルーオンを通して色を交換する
赤クオーク→(赤,反緑)グルーオン放出:渡す色(赤)と受取る色(緑)→緑クオーク
↓
緑クオーク→(赤,反緑)グルーオン吸収:受取る色(赤)と渡す色(緑)→赤クオーク
↓
緑クオーク→(赤,反緑)グルーオン吸収:受取る色(赤)と渡す色(緑)→赤クオーク
グルーオンは与える色ともらう色の組み合わせで8種ある(正方行列3×3=1+8、1:行列の対角成分)
(赤,反青)、(赤,反緑)、(青,反赤)、(青,反緑)、(緑,反赤)、(緑,反青)、(赤,反赤)、(青,反青)、(緑,反緑)
の9種で、最後の3つは、そのうち任意の2つでもう1つを表せるので、グルーオンの色は8種類とされる
(赤,反青)、(赤,反緑)、(青,反赤)、(青,反緑)、(緑,反赤)、(緑,反青)、(赤,反赤)、(青,反青)、(緑,反緑)
の9種で、最後の3つは、そのうち任意の2つでもう1つを表せるので、グルーオンの色は8種類とされる
長さに比例した色の力:ゴムひも状の色力線の閉じ込めでクォークやグルーオンは単独では取り出せない
光子を衝突させてひもが伸びても、核子内からのクォークとの間に、クォークと反クォークの対が生まれ、
ひもが切れて、反クォークと核子内からのクォークが合体して中間子として放出され、クォークは元に戻る
理由:力線のエネルギーよりも新たなクォークと反クォークの対を生成したほうがエネルギー的に安定する
漸近的自由性:クォークの色や色力線(グルーオン)による色の力は離れるほど大きくなり近いほど弱くなる
クォーク同士が近くなると(ひもが弛み)色の力が働かなくなり、近距離内でクォークは自由に動く訳です
光子を衝突させてひもが伸びても、核子内からのクォークとの間に、クォークと反クォークの対が生まれ、
ひもが切れて、反クォークと核子内からのクォークが合体して中間子として放出され、クォークは元に戻る
理由:力線のエネルギーよりも新たなクォークと反クォークの対を生成したほうがエネルギー的に安定する
漸近的自由性:クォークの色や色力線(グルーオン)による色の力は離れるほど大きくなり近いほど弱くなる
クォーク同士が近くなると(ひもが弛み)色の力が働かなくなり、近距離内でクォークは自由に動く訳です
○:u ○ ○ 旧 ○ 新 旧 ○
○ ⇒ ○ ⇒ ○ ◎ ○ ⇒ ○ ◎ ○
●:d ● ● ● u 反u ●
陽子 →→ 対が生まれ 陽子 π0中間子放出
ひもが伸びる ひもが切れる
エネルギーの蓄積 蓄積エネルギーで真空から対が生まれる
○ ⇒ ○ ⇒ ○ ◎ ○ ⇒ ○ ◎ ○
●:d ● ● ● u 反u ●
陽子 →→ 対が生まれ 陽子 π0中間子放出
ひもが伸びる ひもが切れる
エネルギーの蓄積 蓄積エネルギーで真空から対が生まれる
弱い力=「ベータ崩壊を引き起こす力」=「電荷を運ぶ」粒子
弱い相互作用は中性子と陽子を互いに入れ換えることができる
弱い相互作用は電子とニュートリノを互いに入れ換えることができる
弱い相互作用は中性子と陽子を互いに入れ換えることができる
弱い相互作用は電子とニュートリノを互いに入れ換えることができる
|
①電子から出た-電荷の弱ボソンは陽子に入り、電子は-電荷を失いニュートリノになり 陽子は-電荷の弱ボソンを吸収して、電荷が中和され中性子になる:EC
e-──→νe p+──→n W W
②陽子から出た+電荷の弱ボソンが電子に入り、陽子は+電荷を失い中性子になり① ↓W- ② ↓W+ ③ p+──→n+e++νe ④ n ──→p++e-+νe p+──→n e-──→νe (β+崩壊) (β-崩壊) (EC:電子捕獲) (EC:電子捕獲) 電子は+電荷の弱ボソンを吸収して、電荷が中和されニュートリノになる:EC ③陽子が 弱い力の作用で+電荷を失い中性子になり、陽電子とニュートリノを放出:β+ ④中性子が弱い力の作用で+電荷の陽子になり、 電子と反ニュートリノを放出:β- ⑤中性子の周囲にある電子ニュートリノがWボソンを放出して 中性子の中のdクォークがWボソンを吸収してuクォークに変わり、 中性子は陽子になり、電子ニュートリノは電子になる(NC:ニュートリノ捕獲) n(udd)+νe──→W──→ p+(uud)+e- … ④変形、①(EC)の逆反応 ※ニュートリノ・反ニュートリノのスピン ニュートリノと 電子の組は常に左回りにスピンして進む(下向き) 反ニュートリノと陽電子の組は常に右回りにスピンして進む(上向き) |
|
W W
⑥⑦陽子(uud)と中性子(udd)のβ変換は核子のuクォークとdクォークの変換:(u→d)⑥ p+(uud)+e-──→n(udd)+νe ⑦ p+(uud)+μ──→n(udd) + νμ W W ⑧ μ+νe──→νμ+e- ⑧変形: ⑨ μ──→νμ+e-+νe ⑥⑦はクォークとレプトン組の変換、⑧はレプトン組の変換、⑨はミュー粒子変換 ⑥⑦⑧の各組は一つを反転させて左辺から右辺に、右辺から左辺に移す変形が可能 「カミオカンデ」で反ニュートリノを観測、反ニュートリノが弱い力で陽電子に変化
W(反ニュートリノが放出、陽子が吸収)
弱い力は、太陽が燃える際に必要、放射性同位体のベータ崩壊の際にも必要⑥変形=③変形:νe+p+(uud)──→e++n(udd) |
|
中間子の場合、 ⑦式でdクォークを左辺に移して反dクォークに、μ粒子を右辺に移して反μ粒子にする
W
u反d(π+中間子)──→νμ+μ … 弱い力の作用でπ+中間子が反μ粒子に変わる |
|
ストレンジ粒子は弱い力の作用で陽子や中性子になる:ラムダ粒子(uds)→uud/udd |
|
電子・陽電子間、μニュートリノ・電子間に電荷を変えない中性弱ボソン(Z)が作用する |
①光子+陽子(uud) → ラムダ粒子(uds)+K+中間子(su)
②K-中間子(us)+陽子(uud) → π-中間子(ud)+シグマ+粒子(uus)
③ハイパー核 → 通常の原子核
W
ラムダ粒子(uds)+陽子(uud)──→陽子(uud)+中性子(udd)
②K-中間子(us)+陽子(uud) → π-中間子(ud)+シグマ+粒子(uus)
③ハイパー核 → 通常の原子核
W
ラムダ粒子(uds)+陽子(uud)──→陽子(uud)+中性子(udd)
場の古典論:量子の考え方が入っていない場の理論
電場・磁場の理論
場の量子論:波も粒子も量子的な場の振動であり、同じものである
相互作用を媒介するゲージ粒子は場の振動である
重力相互作用を媒介する重力子は未発見であり、
重力相互作用を場の量子論で記述することもできていない
電場・磁場の理論
場の量子論:波も粒子も量子的な場の振動であり、同じものである
相互作用を媒介するゲージ粒子は場の振動である
重力相互作用を媒介する重力子は未発見であり、
重力相互作用を場の量子論で記述することもできていない
マクロの世界の重力
ニュートン力学は、「(重力が十分に大きい)巨視的なスケールで、かつ光速よりも十分遅い速さ
の運動を扱う際の、無矛盾・完結的な近似理論」で「絶対時間」と「絶対空間」を前提とした
アインシュタインの「特殊相対性理論」は、重力の影響を考えない特殊な環境下において、
「時間の進み方や空間の大きさは『絶対的』なものではなく、観測者の置かれた状況によって
変わる『相対的』なものである」とする物理理論です
アインシュタインの一般相対性理論は3次元空間と1次元時間をひとつの4次元時空に統合し、
重力を時空の曲がりの幾何学として記述する
これで多くの重力現象の説明や予言に成功してきた
また2015年には、ブラックホール連星が合体する過程からの重力波が史上初めて直接観測され、
強い重力場での一般相対性理論の検証が一段と進歩した今後、さらに多くの重力波源が観測され、
様々な極限的状況下の一般相対性理論の性質がより明らかになると期待されている
ニュートン力学は、「(重力が十分に大きい)巨視的なスケールで、かつ光速よりも十分遅い速さ
の運動を扱う際の、無矛盾・完結的な近似理論」で「絶対時間」と「絶対空間」を前提とした
アインシュタインの「特殊相対性理論」は、重力の影響を考えない特殊な環境下において、
「時間の進み方や空間の大きさは『絶対的』なものではなく、観測者の置かれた状況によって
変わる『相対的』なものである」とする物理理論です
アインシュタインの一般相対性理論は3次元空間と1次元時間をひとつの4次元時空に統合し、
重力を時空の曲がりの幾何学として記述する
これで多くの重力現象の説明や予言に成功してきた
また2015年には、ブラックホール連星が合体する過程からの重力波が史上初めて直接観測され、
強い重力場での一般相対性理論の検証が一段と進歩した今後、さらに多くの重力波源が観測され、
様々な極限的状況下の一般相対性理論の性質がより明らかになると期待されている
ミクロの世界の重力
実験的には0.01ミリメートルより小さい距離では重力がどのように働くかは我々は知らない
それより小さい距離では重力が我々のまったく予想していなかった振る舞いをしているかもしれない
たとえば小さい距離では隠れた次元があるかもしれない
宇宙初期には全く異なる重力理論が宇宙を支配していたかもしれない
超弦理論やM理論など多くの理論はそのような余剰次元の存在や重力理論の修正を示唆する
余剰次元が宇宙のいたる所に存在していて我々には見えないだけなのかもしれない
余剰次元は直接に見ることはできないが、
高エネルギー実験や宇宙観測から間接的証拠を得られるかもしれない
実験的には0.01ミリメートルより小さい距離では重力がどのように働くかは我々は知らない
それより小さい距離では重力が我々のまったく予想していなかった振る舞いをしているかもしれない
たとえば小さい距離では隠れた次元があるかもしれない
宇宙初期には全く異なる重力理論が宇宙を支配していたかもしれない
超弦理論やM理論など多くの理論はそのような余剰次元の存在や重力理論の修正を示唆する
余剰次元が宇宙のいたる所に存在していて我々には見えないだけなのかもしれない
余剰次元は直接に見ることはできないが、
高エネルギー実験や宇宙観測から間接的証拠を得られるかもしれない
宇宙の大規模構造の重力
数十億光年といったような大きな距離での重力も一般相対性理論とは異なるかもしれない
最近の精密観測は宇宙の膨張が加速していることを明らかにした
これは、もし一般相対性理論が正しければ、
宇宙の70%以上が見えない、しかも圧力が負のエネルギーで満たされていることを意味する
このエネルギーは暗黒エネルギーと呼ばれているが、その正体はまったくわかっていない
アインシュタイン理論を大きな距離で変更して暗黒エネルギーの謎を解明できないか、探究が続く
数十億光年といったような大きな距離での重力も一般相対性理論とは異なるかもしれない
最近の精密観測は宇宙の膨張が加速していることを明らかにした
これは、もし一般相対性理論が正しければ、
宇宙の70%以上が見えない、しかも圧力が負のエネルギーで満たされていることを意味する
このエネルギーは暗黒エネルギーと呼ばれているが、その正体はまったくわかっていない
アインシュタイン理論を大きな距離で変更して暗黒エネルギーの謎を解明できないか、探究が続く
| 力の伝達粒子 | 力の作用先 | 力の種類 | 力の働き | 力の源 |
|---|---|---|---|---|
| 光子 | 原子核・電子 | 電磁気力 | 原子核と電子間の電気力 | 電荷 |
| 中間子 | 陽子・中性子 | 強い力(核力) | 陽子と中性子間の核力 | 色荷(カラー) |
| グルーオン | クォーク | 強い力(色の力) | クォーク同士の色の力 | |
| 弱ボソン | フェルミ粒子 | 弱い力 | 電荷を運ぶ(β変換) | 弱荷=弱アイソスピン |
| 素粒子 | フェルミ粒子 (基本粒子) | 物質粒子 | 第1世代 | 第2世代 | 第3世代 | 電荷 | スピン |
|---|---|---|---|---|---|---|---|
| クォーク (ハドロン構成粒子) | u(アップ) | c(チャーム) | t(トップ) | 2/3 | 1/2 | ||
| d(ダウン) | s(ストレンジ) | b(ボトム) | -1/3 | ||||
| レプトン (軽粒子) | 電子e-,陽電子e+ | ミュー粒子μ± | タウ粒子τ± | ±1 | |||
| 電子ニュートリノνe | ミューニュートリノνμ | タウニュートリノντ | 0 | ||||
| ボース粒子 | 力の伝達粒子 | 電磁相互作用 | 弱い相互作用 | 強い相互作用 | 電荷 | スピン | |
| ゲージ粒子 (力の粒子) | フォトン (γ:光子) | 弱ボソン (W±,Z) | グルーオン (g:膠着子) | (注1) | 1 | ||
| 粒子に質量を与える粒子 | 電荷 | スピン | |||||
| スカラー粒子 | ヒッグス粒子(H) | 0 | 0 | ||||
| 複合粒子 | ハドロン (強粒子) | バリオン (重粒子) | 核子(陽子p,中性子n),デルタ粒子⊿ | (計算) | (注2) | ||
| ハイペロン(ラムダ粒子Λ,シグマ粒子Σ,グザイ粒子Ξ,オメガ粒子Ω) | (計算) | (注3) | |||||
| メソン(中間子) | π,η,ρ,ω,φ,J/ψ,Y,K,B,D | (計算) | (注4) | ||||
| その他 | 原子核,ハイパー核,原子,分子,イオンなど | ||||||
注1:Wボソンは電荷±1、それ以外のゲージ粒子は電荷0
注2:核子はスピン1/2、デルタ粒子はスピン3/2
スピンが3/2ということは、デルタ粒子を構成する3つのクォークは、3つのうち1つが反対の
方向を向いている核子とは異なり、全て同じ方向を向いていることを示している
デルタ粒子は強い力の作用で瞬時に核子に変わる
注3:ハイペロンはストレンジ粒子とも言われ、奇妙さに応じsクォークを1~3個もつ
オメガ粒子はスピン3/2、それ以外のハイペロンはスピン1/2
ハイペロンは弱い力の作用で中間子を放出して、ゆっくりと核子に変わる
注4:擬スカラー中間子はスピン0:π中間子,η中間子など
ベクター中間子はスピン1:ρ中間子,ω中間子,φ中間子,J/ψ中間子,Υ中間子など
K,D,B中間子は荷電の有無や構成クォ―クで擬スカラー中間子またはベクター中間子に属す
上記以外:仮説上の素粒子(重力子など),仮説上の複合粒子,準粒子他多数のハドロンが発見されている
核子や中間子のように核力を感じる物質粒子を「強粒子(ハドロン、hadron)」と総称
核力は感じないものの、電磁気力と弱い力を感じる物質粒子を「軽粒子(レプトン、lepton)」と総称
注2:核子はスピン1/2、デルタ粒子はスピン3/2
スピンが3/2ということは、デルタ粒子を構成する3つのクォークは、3つのうち1つが反対の
方向を向いている核子とは異なり、全て同じ方向を向いていることを示している
デルタ粒子は強い力の作用で瞬時に核子に変わる
注3:ハイペロンはストレンジ粒子とも言われ、奇妙さに応じsクォークを1~3個もつ
オメガ粒子はスピン3/2、それ以外のハイペロンはスピン1/2
ハイペロンは弱い力の作用で中間子を放出して、ゆっくりと核子に変わる
注4:擬スカラー中間子はスピン0:π中間子,η中間子など
ベクター中間子はスピン1:ρ中間子,ω中間子,φ中間子,J/ψ中間子,Υ中間子など
K,D,B中間子は荷電の有無や構成クォ―クで擬スカラー中間子またはベクター中間子に属す
上記以外:仮説上の素粒子(重力子など),仮説上の複合粒子,準粒子他多数のハドロンが発見されている
核子や中間子のように核力を感じる物質粒子を「強粒子(ハドロン、hadron)」と総称
核力は感じないものの、電磁気力と弱い力を感じる物質粒子を「軽粒子(レプトン、lepton)」と総称
○生体の構成:生き物─→器官─→組織─→細胞小器官─→組織化された高分子─→分子や原子
オルガネラ 高分子:タンパク質,核酸,多糖類
オルガネラ 高分子:タンパク質,核酸,多糖類
| / → \ |
| / → \ |
| / → \ |
| / → \ |
| ||||||||||
|
10倍に拡大 | ↑| 10倍に拡大 | ↑↑
【ピンポン玉:水素原子】 ≒ 【地球:ピンポン玉】| || └→1千倍に拡大→┘| | || | └→1万倍に拡大→┘└─────→1万倍に拡大─────→┘ 【原子:原子核】 ≒ 【甲子園球場:1円玉】 | ||||||||||||||||||
原子・分子の世界:電子を動かす電気の力を伝える光子、電子と光子が主役
原子核・素粒子の世界:太陽や宇宙や超高温・高エネルギー現象、究極の粒子
原子核の大きさは原子の1万分の1、原子核の比重は原子の比重の数兆倍程度
原子核・素粒子の世界:太陽や宇宙や超高温・高エネルギー現象、究極の粒子
原子核の大きさは原子の1万分の1、原子核の比重は原子の比重の数兆倍程度
陰極線の発見─┬→電磁波の発見→X線の発見→放射線の発見→原子核の発見
│ (原子にα線照射)
└→電子の発見(陰極線は電子)
陽子の発見→中性子の発見→中間子の発見→ニュートリノの発見│ (原子にα線照射)
└→電子の発見(陰極線は電子)
反陽子の発見→クォークの存在予言と発見→ヒッグス粒子の発見
電子以外の素粒子は20世紀に発見、ヒッグス粒子は2012年に発見
核子は3つのクォークからなる複合粒子、中間子はクォークと反クォークからなる複合粒子
クォーク:((u,d),(c,s),(t,b))、反クォーク:((反u,反d),(反c,反s),(反t,反b))
クォーク:((u,d),(c,s),(t,b))、反クォーク:((反u,反d),(反c,反s),(反t,反b))
素粒子の電荷:(u,c,t)クォーク=2/3、(d,s,b)クォーク=-1/3、(e,μ,τ)=±1、ν=0
ハドロンの電荷:陽子(uud)=1、中性子(udd)=0、デルタ粒子(uuu,uud,udd,ddd)=(2,1,0,-1)、
π+中間子(u反d)=1、π-中間子(d反u)=-1、π0中間子=(u反u)or(d反d)=0
ラムダ粒子(uds)、シグマ粒子(uus,uds,dds)、グザイ粒子(uss,dss)、オメガ粒子(sss)
Λ( 0)、 Σ( 1, 0, -1)、 Ξ( 0, -1)、 Ω( -1)
ハドロンの電荷:陽子(uud)=1、中性子(udd)=0、デルタ粒子(uuu,uud,udd,ddd)=(2,1,0,-1)、
π+中間子(u反d)=1、π-中間子(d反u)=-1、π0中間子=(u反u)or(d反d)=0
ラムダ粒子(uds)、シグマ粒子(uus,uds,dds)、グザイ粒子(uss,dss)、オメガ粒子(sss)
Λ( 0)、 Σ( 1, 0, -1)、 Ξ( 0, -1)、 Ω( -1)
粒子と電荷が反対の反粒子が出会うと中間子や光子を放出して消滅する(クォークと反クォークが消滅する)
中間子はクォークと反クォークからなり、中間子と核子、+中間子と-中間子が別の核子や別の中間子になる
陽子(uud)+π-中間子(d,反u)=中性子(udd)、中性子(udd)+π+中間子(u,反d)=陽子(uud)
陽子(uud)=中性子(udd)+π+中間子(u,反d)、中性子(udd)=陽子(uud)+π-中間子(d,反u)
K+中間子(u,反s)+K-中間子(反u,s)=Φ中間子(s,反s)
バリオン:クォーク3つでできた白色状態、バリオンの反粒子反バリオン:反クォーク3つでできた白色状態
中間子はクォークと反クォークからなり、中間子と核子、+中間子と-中間子が別の核子や別の中間子になる
陽子(uud)+π-中間子(d,反u)=中性子(udd)、中性子(udd)+π+中間子(u,反d)=陽子(uud)
陽子(uud)=中性子(udd)+π+中間子(u,反d)、中性子(udd)=陽子(uud)+π-中間子(d,反u)
K+中間子(u,反s)+K-中間子(反u,s)=Φ中間子(s,反s)
バリオン:クォーク3つでできた白色状態、バリオンの反粒子反バリオン:反クォーク3つでできた白色状態
原子や分子、素粒子などの量子状態を表わす数値として、量子力学で使われる各種の「量子数」がある
量子状態とは電子のような粒子を見分けるための量子数の組み合わせ(=個体識別データ)
電子の量子状態を決める量子数は次の4種類:電子が原子核のまわりを公転する電子軌道を決定する①②③
①主量子数n :軌道の大きさ(原子核・電子間の距離、電子殻:K,L,M,N,…殻)n=1,2,…,7
②方位量子数l:軌道の形 (円・楕円、電子軌道の種類:spdf軌道)l=0,1,2,…,n-1
③磁気量子数m:軌道の向き (軌道の立体的な角度:電子軌道は磁場の中にある)-l≦m≦l
④スピンs :磁気的な性質(電子の自転?回転角運動量?電子の磁気?)+1/2,-1/2
粒子には電荷や性質(内部量子数)の符号が反対の粒子(=反粒子)があるが、区別のない光子などもある
内部量子数(運動量や座標によらない量子数):フレーバー(6種のクォーク),電荷,バリオン数,レプトン数等
2種のクォークは3世代で6種のフレーバー、3種の色の状態、スピンが2種の回り方で計36種類の状態です
量子状態とは電子のような粒子を見分けるための量子数の組み合わせ(=個体識別データ)
電子の量子状態を決める量子数は次の4種類:電子が原子核のまわりを公転する電子軌道を決定する①②③
①主量子数n :軌道の大きさ(原子核・電子間の距離、電子殻:K,L,M,N,…殻)n=1,2,…,7
②方位量子数l:軌道の形 (円・楕円、電子軌道の種類:spdf軌道)l=0,1,2,…,n-1
③磁気量子数m:軌道の向き (軌道の立体的な角度:電子軌道は磁場の中にある)-l≦m≦l
④スピンs :磁気的な性質(電子の自転?回転角運動量?電子の磁気?)+1/2,-1/2
粒子には電荷や性質(内部量子数)の符号が反対の粒子(=反粒子)があるが、区別のない光子などもある
内部量子数(運動量や座標によらない量子数):フレーバー(6種のクォーク),電荷,バリオン数,レプトン数等
2種のクォークは3世代で6種のフレーバー、3種の色の状態、スピンが2種の回り方で計36種類の状態です
フェルミ粒子:クォーク,レプトン,核子(陽子,中性子) … スピンが半整数になるのがフェルミオン
ボース粒子 :ゲージ粒子,重力子,ヒッグス粒子,中間子 … スピンが整数になるのがボソン
パウリの排他律:ボース粒子は重ねられるけれどフェルミ粒子は重ねられない(スピンが性格を決める)
・ボース粒子 :2個以上の粒子が同じ状態をとれる (同じ位置に重ねられる、 空間的大きさがない)
・フェルミ粒子:一つの状態を1個の粒子しかとれない(同じ位置に重ねられない、空間的大きさがある)
ボース粒子 :ゲージ粒子,重力子,ヒッグス粒子,中間子 … スピンが整数になるのがボソン
パウリの排他律:ボース粒子は重ねられるけれどフェルミ粒子は重ねられない(スピンが性格を決める)
・ボース粒子 :2個以上の粒子が同じ状態をとれる (同じ位置に重ねられる、 空間的大きさがない)
・フェルミ粒子:一つの状態を1個の粒子しかとれない(同じ位置に重ねられない、空間的大きさがある)
ボース粒子は極低温でボース・アインシュタイン凝縮(BEC理論)を起こす
ある転移温度以下で巨視的な数のボース粒子がある1つの1粒子状態に落ち込む相転移現象
ボース粒子であるヘリウム4はその凝縮を起源とする超流動状態(粘性ゼロ)を示す
フェルミ粒子2個は疑似ボース粒子としてボース・アインシュタイン凝縮を起こす
電子の対はその凝縮を起源とする超伝導状態(電気抵抗ゼロ)を示す … 電荷を持った粒子の超流動
ヘリウム3の対はその凝縮を起源とする超流動状態を示す
ある転移温度以下で巨視的な数のボース粒子がある1つの1粒子状態に落ち込む相転移現象
ボース粒子であるヘリウム4はその凝縮を起源とする超流動状態(粘性ゼロ)を示す
フェルミ粒子2個は疑似ボース粒子としてボース・アインシュタイン凝縮を起こす
電子の対はその凝縮を起源とする超伝導状態(電気抵抗ゼロ)を示す … 電荷を持った粒子の超流動
ヘリウム3の対はその凝縮を起源とする超流動状態を示す
素粒子のスピンは大きさと向きをもち、クォークのスピンの大きさは1/2,向きは右回りが(+),左回りが(-)
核子のスピン:核子内の3つのクォークのうち2つは右回り(+)、1つは左回り(-)で、全体で右回り(+)の
スピン1/2です:+1/2+1/2-1/2=+1/2 … uクォークが+、dクォークが-ではなく、各々+か-がある
ラムダ粒子はuクォークとdクォークのスピンが逆で打ち消しあいラムダ粒子のスピンはsクォークのスピンです
デルタ粒子は3つのクォークのスピンの向きが同じ(左/右)3/2
核子は特定のエネルギーの光子を吸収して左回りの1つが右回りになりスピンの向きが同じデルタ粒子になる
電荷がありスピンしているクォークはミニ磁石で、クォーク磁石が3つ合わさって核子の磁石になる
核子のスピン:核子内の3つのクォークのうち2つは右回り(+)、1つは左回り(-)で、全体で右回り(+)の
スピン1/2です:+1/2+1/2-1/2=+1/2 … uクォークが+、dクォークが-ではなく、各々+か-がある
ラムダ粒子はuクォークとdクォークのスピンが逆で打ち消しあいラムダ粒子のスピンはsクォークのスピンです
デルタ粒子は3つのクォークのスピンの向きが同じ(左/右)3/2
核子は特定のエネルギーの光子を吸収して左回りの1つが右回りになりスピンの向きが同じデルタ粒子になる
電荷がありスピンしているクォークはミニ磁石で、クォーク磁石が3つ合わさって核子の磁石になる
電子のスピンは上向きスピンと下向きスピンの2種類
パウリの排他原理:ひとつの電子軌道に入れる電子は2個までで、そのスピンは逆でなければならない
同じ量子状態の共有を禁ずるのがパウリの排他原理
最低ひとつの量子数は違っていなければならない、軌道が同じならスピンは異なってなくてはならない
ひとつの電子軌道に複数の電子が入るにはスピンが違っていなければならないしスピンは2通りしかない
ひとつの電子軌道に入れる電子は互いにスピンが逆のふたつだけ
電荷がありスピンしている電子はミニ磁石です(電流と磁界の向きはアンペールの右ねじの法則に従う)
上向きスピンは下から見て右回り(+)でN極が上、下向きスピンは下から見て左回り(-)でN極が下
アンペール、ビオ・サバール ファラデー、フレミング
パウリの排他原理:ひとつの電子軌道に入れる電子は2個までで、そのスピンは逆でなければならない
同じ量子状態の共有を禁ずるのがパウリの排他原理
最低ひとつの量子数は違っていなければならない、軌道が同じならスピンは異なってなくてはならない
ひとつの電子軌道に複数の電子が入るにはスピンが違っていなければならないしスピンは2通りしかない
ひとつの電子軌道に入れる電子は互いにスピンが逆のふたつだけ
電荷がありスピンしている電子はミニ磁石です(電流と磁界の向きはアンペールの右ねじの法則に従う)
上向きスピンは下から見て右回り(+)でN極が上、下向きスピンは下から見て左回り(-)でN極が下
N S
→ ▲ ← △
+1/2 ↑○↓ ◎ -1/2 ↓○↑ ◎
← ▽ → ▼
S N
「電流と磁界に関する法則」と「電磁誘導に関する法則」は別ものなので混同しないようにしましょう→ ▲ ← △
+1/2 ↑○↓ ◎ -1/2 ↓○↑ ◎
← ▽ → ▼
S N
アンペール、ビオ・サバール ファラデー、フレミング
デルタ++粒子は3つのクォークが、同じ電荷+2/3の状態、同じuのフレーバーの状態、同じスピンの向きで、
同じ場所にあり、パウリの排他原理に違反しないためには3つの異なる色の状態が必要になる
クォークの新しい3種の状態として、赤・青・緑の3原色が考えられた
オメガ-粒子内の3つのクォークも同じsフレーバー、同じ-1/3電荷ですが、3つの異なる色の状態をもつ
核子・デルタ粒子・ハイペロンなどの重粒子は3つのクォークが3つの色の衣を持ち、中間子はクォーク
・反クォークが2つの補色関係の色の衣をもつ(クォークや中間子の色の衣は時々衣を交換する)
クォークの閉じ込め:クォークが単独では存在しないのは、色は単色では存在できず、無色の粒子だけが
単独で存在できる事や、単独の分数電荷では存在できず、整数電荷だけが単独で存在できる事を意味する
同じ場所にあり、パウリの排他原理に違反しないためには3つの異なる色の状態が必要になる
クォークの新しい3種の状態として、赤・青・緑の3原色が考えられた
オメガ-粒子内の3つのクォークも同じsフレーバー、同じ-1/3電荷ですが、3つの異なる色の状態をもつ
核子・デルタ粒子・ハイペロンなどの重粒子は3つのクォークが3つの色の衣を持ち、中間子はクォーク
・反クォークが2つの補色関係の色の衣をもつ(クォークや中間子の色の衣は時々衣を交換する)
クォークの閉じ込め:クォークが単独では存在しないのは、色は単色では存在できず、無色の粒子だけが
単独で存在できる事や、単独の分数電荷では存在できず、整数電荷だけが単独で存在できる事を意味する
フェルミ粒子:軽い順に第1世代,第2世代,第3世代と呼ぶが、現在の原子の構成要素は第1世代のみ
他の世代は、宇宙創成時に存在したが現在の地球にはなく、加速粒子を衝突させて、人工的に作られる
ミュー粒子は、宇宙線が大気中の原子と衝突して生成され地表に到達する2次宇宙線として観測される
世代が高い粒子は、それより前の世代の対応する粒子よりも大きな質量を持つ … 質量階層
各世代間は、粒子の質量のみ異なり、全ての基本相互作用および量子数は同一
他の世代は、宇宙創成時に存在したが現在の地球にはなく、加速粒子を衝突させて、人工的に作られる
ミュー粒子は、宇宙線が大気中の原子と衝突して生成され地表に到達する2次宇宙線として観測される
世代が高い粒子は、それより前の世代の対応する粒子よりも大きな質量を持つ … 質量階層
各世代間は、粒子の質量のみ異なり、全ての基本相互作用および量子数は同一
核子と高エネルギーの中間子や光子の衝突でハイペロン(ストレンジ粒子)が作られる
ラムダ粒子(uds),シグマ粒子(uus,uds,dds),グザイ粒子(uss,dss),オメガ粒子(sss)
核子内の3つのクォークのうち1個~3個がsクォークであるのがハイペロンです
ハイペロンを含む原子核をハイパー核と言い、ハイパー核内のハイペロンは弱い力
の作用で陽子や中性子に変わり、ハイパー核は通常の原子核に戻る
ラムダ粒子(uds),シグマ粒子(uus,uds,dds),グザイ粒子(uss,dss),オメガ粒子(sss)
核子内の3つのクォークのうち1個~3個がsクォークであるのがハイペロンです
ハイペロンを含む原子核をハイパー核と言い、ハイパー核内のハイペロンは弱い力
の作用で陽子や中性子に変わり、ハイパー核は通常の原子核に戻る
レプトンの各世代は弱い力の作用で±W弱ボソンを交換して変換する(ただし同世代の中に限る)
電荷を持つ粒子と電荷を持たない粒子間で双方向に変換する(e⇔νe、μ⇔νμ、τ⇔ντ)
μ粒子は百万分の22秒の寿命でμニュートリノに変わり、電子と反電子ニュートリノを放出する
τ粒子は10兆分の3秒の寿命でτニュートリノに変わり、電子と反電子ニュートリノを放出する
か、μ粒子と反μニュートリノを放出する
電荷を持つ粒子と電荷を持たない粒子間で双方向に変換する(e⇔νe、μ⇔νμ、τ⇔ντ)
μ粒子は百万分の22秒の寿命でμニュートリノに変わり、電子と反電子ニュートリノを放出する
τ粒子は10兆分の3秒の寿命でτニュートリノに変わり、電子と反電子ニュートリノを放出する
か、μ粒子と反μニュートリノを放出する
ニュートリノには電子ニュートリノ、μニュートリノ、τニュートリノの3種類があります
これは「フレーバー」による分類です … 電子型νe、ミュー型νμ、タウ型ντ
ニュートリノは物質に当たって弱い相互作用を受けると、荷電レプトンに変わります
・電子に変わるニュートリノを電子ニュートリノ
・μ粒子に変わるニュートリノをμニュートリノ
・τ粒子に変わるニュートリノをτニュートリノ
3種類の荷電レプトン(e,μ,τ) : 質量でのみ識別可能
3種類のニュートリノ : 物質と相互作用することによって識別可能=フレーバー
一方、ニュートリノは「質量」という分類で分けることもできます
m1,m2,m3という3つの異なる質量を持つ、
ニュートリノ1、ニュートリノ2、ニュートリノ3の3種類です … ν1、ν2、ν3
ニュートリノ混合:ニュートリノν1、ν2、ν3の混合
νe=[ν1>ν2>ν3]、νμ=[ν1<ν2≒ν3]、ντ=[ν1<ν2<ν3]
ニュートリノは、異なる質量を持つ複数の可能性が重ね合わさった状態
これは「フレーバー」による分類です … 電子型νe、ミュー型νμ、タウ型ντ
ニュートリノは物質に当たって弱い相互作用を受けると、荷電レプトンに変わります
・電子に変わるニュートリノを電子ニュートリノ
・μ粒子に変わるニュートリノをμニュートリノ
・τ粒子に変わるニュートリノをτニュートリノ
3種類の荷電レプトン(e,μ,τ) : 質量でのみ識別可能
3種類のニュートリノ : 物質と相互作用することによって識別可能=フレーバー
一方、ニュートリノは「質量」という分類で分けることもできます
m1,m2,m3という3つの異なる質量を持つ、
ニュートリノ1、ニュートリノ2、ニュートリノ3の3種類です … ν1、ν2、ν3
ニュートリノ混合:ニュートリノν1、ν2、ν3の混合
νe=[ν1>ν2>ν3]、νμ=[ν1<ν2≒ν3]、ντ=[ν1<ν2<ν3]
ニュートリノは、異なる質量を持つ複数の可能性が重ね合わさった状態
①原子炉内の核分裂の際の放射性核のベータ変換、②高エネルギー陽子加速器で生成(下図)
③宇宙、超新星、太陽、大気、地球の裏側、人体からやってくる
電気を帯びずよく透過する、弱い力だけが作用する、電子の100万分の1以下の質量
現時点では、左巻きらせん運動のニュートリノと、右巻きらせん運動の反ニュートリノ
しか見つかっていない(他の素粒子は、粒子は左右あり、反粒子も左右ある)
↑ ↑
π+中間子→μ+粒子→e+
↑ ↓
500MeV │ νe
②高エネルギー陽子→Be原子核
│ νe
↓ ↑
π-中間子→μ-粒子→e-
↓ ↓
νμ νμ
③太陽風陽子の場合→大気中の窒素や酸素→…ミュー粒子、電子、ニュートリノ
③宇宙、超新星、太陽、大気、地球の裏側、人体からやってくる
電気を帯びずよく透過する、弱い力だけが作用する、電子の100万分の1以下の質量
現時点では、左巻きらせん運動のニュートリノと、右巻きらせん運動の反ニュートリノ
しか見つかっていない(他の素粒子は、粒子は左右あり、反粒子も左右ある)
①核反応パイ中間子の変換によるニュートリノ生成
νμ νμ↑ ↑
π+中間子→μ+粒子→e+
↑ ↓
500MeV │ νe
②高エネルギー陽子→Be原子核
│ νe
↓ ↑
π-中間子→μ-粒子→e-
↓ ↓
νμ νμ
③太陽風陽子の場合→大気中の窒素や酸素→…ミュー粒子、電子、ニュートリノ
ニュートリノは、「粒子」であると同時に「波」としての性質を持ちます
それぞれ異なる質量を持つニュートリノν1、ν2、ν3は、
それぞれ異なる振動数を持つ「波」として空間を伝播します
ニュートリノのフレーバーは、3種類の質量の決まった「波」の「重ね合わせ」(合成波)となり、
ニュートリノが空間を飛ぶ間に合成波の位相が変化し、フレーバーの種類(型)が移り変わります
νe⇔νμ、νμ⇔ντ、ντ⇔νe この現象をニュートリノ振動と呼びます
ニュートリノが質量を持ち、ゼロではないニュートリノ混合があるときに起こる現象です
質量がわずかに異なる3種類のニュートリノν1、ν2、ν3は、それぞれ異なる速度で飛行する
これを飛行経路のある場所で観測すると、元々作られた型とは異なるタイプのニュートリノに、
ある確率で変わってしまう … ニュートリノの世代交代(移動)が起こる
ニュートリノ振動が起きる割合はν1、ν2、ν3の質量の二乗の差と飛行距離に比例し、
ニュートリノが持つエネルギーに反比例する
ニュートリノが反応した時に対になっている、電子やμ粒子、τ粒子を観測し、
ニュートリノの型を判別することで型の変化がわかります(型:フレーバーによる分類)
それぞれ異なる質量を持つニュートリノν1、ν2、ν3は、
それぞれ異なる振動数を持つ「波」として空間を伝播します
ニュートリノのフレーバーは、3種類の質量の決まった「波」の「重ね合わせ」(合成波)となり、
ニュートリノが空間を飛ぶ間に合成波の位相が変化し、フレーバーの種類(型)が移り変わります
νe⇔νμ、νμ⇔ντ、ντ⇔νe この現象をニュートリノ振動と呼びます
ニュートリノが質量を持ち、ゼロではないニュートリノ混合があるときに起こる現象です
質量がわずかに異なる3種類のニュートリノν1、ν2、ν3は、それぞれ異なる速度で飛行する
これを飛行経路のある場所で観測すると、元々作られた型とは異なるタイプのニュートリノに、
ある確率で変わってしまう … ニュートリノの世代交代(移動)が起こる
ニュートリノ振動が起きる割合はν1、ν2、ν3の質量の二乗の差と飛行距離に比例し、
ニュートリノが持つエネルギーに反比例する
ニュートリノが反応した時に対になっている、電子やμ粒子、τ粒子を観測し、
ニュートリノの型を判別することで型の変化がわかります(型:フレーバーによる分類)
| 素粒子・複合粒子 | 第1世代 | ||
|---|---|---|---|
| 素粒子 | フェルミ粒子 (基本粒子) | クォーク (ハドロン構成粒子) | u(アップ) |
| d(ダウン) | |||
| レプトン (軽粒子) | 電子e-,陽電子e+ | ||
| 電子ニュートリノνe | |||
| ボース粒子 | ゲージ粒子 (力の粒子) | 光子,弱ボソン(W±,Z) グルーオン(g:膠着子) | |
| スカラー粒子 | ヒッグス粒子(H) | ||
| 複合粒子 | ハドロン (強粒子) | バリオン (重粒子) | 核子(陽子uud,中性子udd) デルタ粒子(uuu,uud,udd,ddd) |
| メソン (中間子) | π+ ,π- ,π0 u反d,d反u,(u反u)or(d反d) | ||
| その他 | 原子核 | ||
フェルミ粒子:クォーク,レプトン,核子(陽子,中性子)
ボース粒子 :ゲージ粒子,重力子,ヒッグス粒子,中間子
量子数 :粒子の量子状態を表わす数値(電荷やスピン)
ボース粒子 :ゲージ粒子,重力子,ヒッグス粒子,中間子
量子数 :粒子の量子状態を表わす数値(電荷やスピン)
二次宇宙線 :ミュー粒子は50万分の1秒の寿命ですが特殊相対性理論で10倍延びて地表に到達する
現存する核子を構成するクォークは第1世代のみで、現存する原子を構成する電子も第1世代である
他の世代のクォークやレプトンは宇宙創成時には存在したが現在の地球にはなく、人工的に作られる
物質の占める割合は多くなく宇宙の全質量の大部分は未知の物質で見えないので暗黒物質と呼ばれる
ニュートリノはいまだに謎の多い粒子でニュートリノの正体と暗黒物質の探索が活発に行われている
現存する核子を構成するクォークは第1世代のみで、現存する原子を構成する電子も第1世代である
他の世代のクォークやレプトンは宇宙創成時には存在したが現在の地球にはなく、人工的に作られる
物質の占める割合は多くなく宇宙の全質量の大部分は未知の物質で見えないので暗黒物質と呼ばれる
ニュートリノはいまだに謎の多い粒子でニュートリノの正体と暗黒物質の探索が活発に行われている
| 種類 | 放射線 | |
|---|---|---|
| 粒子線 | アルファ線 | ヘリウム原子核、電荷:+2 |
| 放射性原子核はアルファ変換で陽子2個中性子2個減り質量数4少ない原子核になる | ||
| ベータ線 | 電子・陽電子、電荷:±1 | |
| 電子が出るベータ変換では、放射性原子核の中性子が陽子になる | ||
| 陽電子が出るベータ変換では、放射性原子核の陽子が中性子になる | ||
| 電子や陽電子が出るベータ変換では、もう一つの粒子ニュートリノが放出される | ||
| 陽子線 | 陽子線:水素の原子核、重陽子線:重水素の原子核、電荷:+1 | |
| 人工的に原子核反応を起こす加速器等から作られる | ||
| 中性子線 | 核分裂などの際に核外に飛び出す中性子、電荷:0 | |
| 原子炉や加速器等から作られる、ベータ壊変して陽子になる | ||
| 電磁波 | ガンマ線 | 光子、波長:10-11m以下 |
| 励起状態の原子核が励起エネルギー分のガンマ線を放出して基底状態になる | ||
| X線 | 光子、波長:10-12~10-8m | |
| 特別なベータ変換では、陽電子放出の代わりに原子核の内側の軌道の電子を吸収して | ||
| 外側の軌道の電子が内側の軌道に移り、軌道差のエネルギーのX線が放出される | ||
放射性変換:アルファ変換、電子/陽電子/X線を放出するベータ変換、ガンマ線放出
電離放射線:荷電粒子線、非荷電粒子線、3千兆ヘルツ以上の電磁波(ガンマ線、X線)
非電離放射線:3千兆ヘルツ以下の電磁波(紫外線の一部、可視光線、赤外線、マイクロ波、電波)
放射線検出器:霧箱によって荷電粒子(アルファ線、ベータ線、宇宙線)の飛跡を見ることができる
放射線測定器:GM管式(ベータ線)
シンチレーション式(ガンマ線、X線)測定器によってはアルファ線も測定可能
電離箱式(ガンマ線、X線)
電離放射線:荷電粒子線、非荷電粒子線、3千兆ヘルツ以上の電磁波(ガンマ線、X線)
非電離放射線:3千兆ヘルツ以下の電磁波(紫外線の一部、可視光線、赤外線、マイクロ波、電波)
放射線検出器:霧箱によって荷電粒子(アルファ線、ベータ線、宇宙線)の飛跡を見ることができる
放射線測定器:GM管式(ベータ線)
シンチレーション式(ガンマ線、X線)測定器によってはアルファ線も測定可能
電離箱式(ガンマ線、X線)
| 電磁波 | 波長 | 主な利用 |
|---|---|---|
| ガンマ線 | 10-10~10-12cm | 科学観測機器 |
| X線 | 10-7cm | 医療機器 |
| 紫外線 | 10-6cm | 殺菌灯 |
| 可視光線 | 10-5cm | 光学機器 |
| 赤外線 | 10-3cm | 赤外線ヒーター |
| サブミリ波 | 0.1cm | 光通信システム |
| 電波 | 102~104cm | 携帯電話・TV・ラジオ・飛行機の通信 |
放射線:天然放射性物質や人工放射性物質からの放射線、宇宙線
放射線の利用:X線撮影、CT検査、重イオンビーム、ガンマナイフ、PET(陽電子放出断層撮影)
ラジオアイソトープ(RI)= 原子炉内の中性子を材料に照射して作られる放射性物質
天然にも存在し、放射性同位元素とも呼ばれ、医療用のほかに、工業・農業分野でも使われている
核医学検査 = 放射線を放出するRIを含んだ薬を使って、ガンマカメラで体内の状態を調べる検査です
①安定な原子核に外から高エネルギー粒子を入射し、原子核反応が起こると放射性核になることがある
②安定な原子核に外から中性子を入射・吸収させると、放射性核ができる
③安定な原子核に高エネルギー陽子を当てて、中の中性子を弾き飛ばすと放射性核ができる
不安定な放射性核は放射線を放出して、静止質量エネルギーの低い原子核(安定核)に変わる
原子力エネルギー:ウラニウムやトリウムの核分裂の際の放出エネルギー … 原子力発電で利用中
核融合エネルギー:重水素や三重水素などの核融合の際の放出エネルギー
数百万~1千万eV:電子線(ベータ線)、陽子線、アルファ線、ガンマ線
1千億eV:宇宙線(ミュー粒子)、化学反応は数eVで放射線エネルギーの百万分の1
放射性変換前後の質量差:E=mc2 … これが放射線エネルギーが高い理由
核分裂の連鎖反応:ウラニウムが中性子を吸収して核分裂が起こり中性子を放出する
核燃料の燃焼制御:原子炉では中性子の減速材や中性子を吸収する制御棒が使われる
核融合の実用化:1億度位の超高温のプラズマをある時間維持し核融合の持続が必要
放射線の利用:X線撮影、CT検査、重イオンビーム、ガンマナイフ、PET(陽電子放出断層撮影)
ラジオアイソトープ(RI)= 原子炉内の中性子を材料に照射して作られる放射性物質
天然にも存在し、放射性同位元素とも呼ばれ、医療用のほかに、工業・農業分野でも使われている
核医学検査 = 放射線を放出するRIを含んだ薬を使って、ガンマカメラで体内の状態を調べる検査です
原子核内の核子の軌道運動→高エネルギー粒子を原子核に衝突させると、
高エネルギー粒子のエネルギーの大きさに応じて下記のいずれかが起こる
原子核の振動→原子核の回転→原子核反応→高温核の生成と核子の蒸発→
沢山の核子が蒸発、終には原子核は核子や小さな原子核にばらばらになる
高エネルギー粒子のエネルギーの大きさに応じて下記のいずれかが起こる
原子核の振動→原子核の回転→原子核反応→高温核の生成と核子の蒸発→
沢山の核子が蒸発、終には原子核は核子や小さな原子核にばらばらになる
①安定な原子核に外から高エネルギー粒子を入射し、原子核反応が起こると放射性核になることがある
②安定な原子核に外から中性子を入射・吸収させると、放射性核ができる
③安定な原子核に高エネルギー陽子を当てて、中の中性子を弾き飛ばすと放射性核ができる
不安定な放射性核は放射線を放出して、静止質量エネルギーの低い原子核(安定核)に変わる
|
宇宙線由来の放射線:中性子→大気中のN14→放射性同位体C14+H、C14+О2…→C14О2
炭素同位体年代測定:遺物や化石に含まれる炭素14(放射性同位体C14)の量を測定する↓放射性崩壊 (半減期5730±40年)→N14+e-+νe
炭素14の中性子1個が弱ボソンの作用で陽子に変わり、窒素14に変わる = β-崩壊
中性子中のdクォークが弱ボソンを放出してuクォークになる 弱ボソンはすぐにこわれて電子と反電子ニュートリノに変わる 炭素14=陽子6個+中性子8個 窒素14=陽子7個+中性子7個 |
原子力エネルギー:ウラニウムやトリウムの核分裂の際の放出エネルギー … 原子力発電で利用中
核融合エネルギー:重水素や三重水素などの核融合の際の放出エネルギー
数百万~1千万eV:電子線(ベータ線)、陽子線、アルファ線、ガンマ線
1千億eV:宇宙線(ミュー粒子)、化学反応は数eVで放射線エネルギーの百万分の1
放射性変換前後の質量差:E=mc2 … これが放射線エネルギーが高い理由
核分裂の連鎖反応:ウラニウムが中性子を吸収して核分裂が起こり中性子を放出する
核燃料の燃焼制御:原子炉では中性子の減速材や中性子を吸収する制御棒が使われる
核融合の実用化:1億度位の超高温のプラズマをある時間維持し核融合の持続が必要
|
中性子(点火剤)→U235核分裂→中性子+U238(天然核燃料)→U239→β変換→Np239→β変換→Pu239 →I131,Cs137,Sr90…,Xe133→β変換→Cs133+中性子→Cs134 Pu239は人工核燃料になるが、一部は中性子を吸収してPu240になり、一部は中性子により核分裂する 核分裂による静止質量エネルギーの変化はウラニウム核あたり0.01%、エネルギーにして2億電子ボルト 半減期が数万年以上の放射能をもつ長寿命核種MA(Np,Am,Cm)の群分離・核変換技術の開発が求められている 資源量の多いトリウムによるトリウム燃料サイクルは、MAの生成量が小さい利点があるが課題も多い |
|
500万eVの放射線の速さと透過する厚さ 1000万eVの放射線の防御 アルファ線 :光速の5%、 アルミホイル0.002cmで止まる 銅箔0.03mm 陽子線 :光速の10%、 アルミ板0.02cmで止まる 銅箔0.25mm ベータ線(電子):光速の99.5%、アルミ板1cmで止まる 銅板6mm ガンマ線(光子):光速、 アルミレンガ20cmで1/4に減少 銅ブロック10cmで1/10,15cmで2/100に減少 中性子線 :光速の4.67%、 数十cm~数mのコンクリートで減らす |
|
宇宙線 :ほぼ光速に近い速さ、 1000億eV、地下250m |
ガンマ線:物質中の電子に何回か衝突すると吸収されて消滅する(一部又は全部が電子のエネルギーになる)
光子は電子に衝突して、散乱、吸収、電子・陽電子対に変換される
中性子線:物質中の原子核に衝突して吸収されて消滅する
放射性物質は、壊変(崩壊)を繰り返し、最終的に安定した物質へ変化すると放射線を放出しなくなります
原子核の壊変には、規則性があり、放射能の量はある一定の時間が経過すると半分になり、更にその同じ
一定の時間が経過するとまたその半分になります … (1→1/2→1/4→1/8…)一定の時間:半減期は同じ
放射線の強度(ベクレル):放射性核数×変換率(%/年)、平均寿命=1/変換率、半減期=平均寿命の約70%
原子核の壊変には、規則性があり、放射能の量はある一定の時間が経過すると半分になり、更にその同じ
一定の時間が経過するとまたその半分になります … (1→1/2→1/4→1/8…)一定の時間:半減期は同じ
放射線の強度(ベクレル):放射性核数×変換率(%/年)、平均寿命=1/変換率、半減期=平均寿命の約70%
|
物理学的半減期:壊変によって、始めの原子核の数が半分になるまでの時間を半減期といいます 半減期は、放射性物質によって違い、数秒のものから100億年を超えるものまであります 生物学的半減期:体内に取り込まれた放射性物質の量が代謝や排泄により体の外へ排出されて半分になる までの時間 実効半減期 :物理学的半減期と生物学的半減期の両方を考慮したもの |
| 放射性物質 | 物理学的半減期 | 放射性物質 | 物理学的半減期 |
|---|---|---|---|
| クリプトン90 | 32.3秒 | クリプトン85 | 11 |
| キセノン138 | 14.1分 | トリチウム | 12.3 |
| フッ素18 | 109.8分 | ストロンチウム90 | 28.8 |
| ナトリウム24 | 15時間 | セシウム137 | 30.0 |
| ラドン222 | 3.8日 | ラジウム226 | 1600 |
| キセノン133 | 5.2日 | 炭素14 | 5700 |
| ヨウ素131 | 8.04日 | プルトニウム239 | 2.4万 |
| ストロンチウム89 | 50.5日 | ウラン235 | 7.0億 |
| ポロニウム210 | 138.4日 | カリウム40 | 13億 |
| セシウム134 | 2.1 | ウラン238 | 45億 |
| コバルト60 | 5.27 | トリウム232 | 141億 |
| 系列:放射性原子核から安定な原子核になるまで次々に核種が変化しながら壊変する | |||
| 非系列:系列を作らず放射性原子核から直接(1回の壊変で)安定な原子核になる | |||
|
ベクレル (Bq):1秒間に壊変(崩壊)する原子核の数で、放射能を表す単位、Bq/ℓ、Bq/kg シーベルト(Sv):体が受けた放射線による影響の度合いを表す単位、ミリSv、μSv グレイ (Gy):放射線のエネルギーが物質や人体の組織に吸収された量を表す単位、ナノGy、μGy |
※原子爆弾の放出したエネルギーの50%は爆風に、 35%は熱線に、15%は放射線となりました
|
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 族→ 周期 |
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 |
| 1 | 1 H 水素 | 2 He ヘリウム | ||||||||||||||||
| 2 | 3 Li リチウム |
4 Be ベリリウム |
上段:原子番号(陽子数)、下段:日本名 中段:元素記号(文字色は常温の相) |
5 B ホウ素 |
6 C 炭素 |
7 N 窒素 |
8 O 酸素 |
9 F フッ素 |
10 Ne ネオン | |||||||||
| 3 | 11 Na ナトリウム |
12 Mg マグネシウム | 13 Al アルミニウム |
14 Si ケイ素 |
15 P リン |
16 S 硫黄 |
17 Cl 塩素 |
18 Ar アルゴン | ||||||||||
| 4 | 19 K カリウム |
20 Ca カルシウム |
21 Sc スカンジウム |
22 Ti チタン |
23 V バナジウム |
24 Cr クロム |
25 Mn マンガン |
26 Fe 鉄 |
27 Co コバルト |
28 Ni ニッケル |
29 Cu 銅 |
30 Zn 亜鉛 |
31 Ga ガリウム |
32 Ge ゲルマニウム |
33 As ヒ素 |
34 Se セレン |
35 Br 臭素 |
36 Kr クリプトン |
| 5 | 37 Rb ルビジウム |
38 Sr ストロンチウム |
39 Y イットリウム |
40 Zr ジルコニウム |
41 Nb ニオブ |
42 Mo モリブデン |
43 Tc テクネチウム |
44 Ru ルテニウム |
45 Rh ロジウム |
46 Pd パラジウム |
47 Ag 銀 |
48 Cd カドミウム |
49 In インジウム |
50 Sn スズ |
51 Sb アンチモン |
52 Te テルル |
53 I ヨウ素 |
54 Xe キセノン |
| 6 | 55 Cs セシウム |
56 Ba バリウム |
ランタ ノイド |
72 Hf ハフニウム |
73 Ta タンタル |
74 W タングステン |
75 Re レニウム |
76 Os オスミウム |
77 Ir イリジウム |
78 Pt 白金 |
79 Au 金 |
80 Hg 水銀 |
81 Tl タリウム |
82 Pb 鉛 |
83 Bi ビスマス |
84 Po ポロニウム |
85 At アスタチン |
86 Rn ラドン |
| 7 | 87 Fr フランシウム |
88 Ra ラジウム |
アクチ ノイド |
104 Rf ラザホージウム |
105 Db ドブニウム |
106 Sg シーボーギウム |
107 Bh ボーリウム |
108 Hs ハッシウム |
109 Mt マイトネリウム |
110 Ds ダームスタチウム |
111 Rg レントゲニウム |
112 Cn コペルニシウム |
113 Nh ニホニウム |
114 Fl フレロビウム |
115 Mc モスコビウム |
116 Lv リバモリウム |
117 Ts テネシン |
118 Og オガネソン |
上記周期表は「長周期型周期表」と呼ばれるもので、「ランタノイド(57番〜71番元素)」
や「アクチノイド(89番〜103番元素)」といった元素群が、表の下側に置かれています
一方、超長周期型の周期表では、ランタノイドやアクチノイドも一つの表の中におさめます
つまり、長周期型と超長周期型は、元素の配置の仕方がちがうだけで、内容は同じものです
や「アクチノイド(89番〜103番元素)」といった元素群が、表の下側に置かれています
一方、超長周期型の周期表では、ランタノイドやアクチノイドも一つの表の中におさめます
つまり、長周期型と超長周期型は、元素の配置の仕方がちがうだけで、内容は同じものです
| ランタ ノイド |
57 La ランタン |
58 Ce セリウム |
59 Pr プラセオジム |
60 Nd ネオジム |
61 Pm プロメチウム |
62 Sm サマリウム |
63 Eu ユウロピウム |
64 Gd ガドリニウム |
65 Tb テルビウム |
66 Dy ジスプロシウム |
67 Ho ホルミウム |
68 Er エルビウム |
69 Tm ツリウム |
70 Yb イッテルビウム |
71 Lu ルテチウム |
| アクチ ノイド |
89 Ac アクチニウム |
90 Th トリウム |
91 Pa プロトアクチニウム |
92 U ウラン |
93 Np ネプツニウム |
94 Pu プルトニウム |
95 Am アメリシウム |
96 Cm キュリウム |
97 Bk バークリウム |
98 Cf カリホルニウム |
99 Es アインスタイニウム |
100 Fm フェルミウム |
101 Md メンデレビウム |
102 No ノーベリウム |
103 Lr ローレンシウム |
背景の色=非金属、アルカリ金属、アルカリ土類金属、遷移金属、ハロゲン、貴ガス、典型金属
金属元素:金属元素、非金属元素:金属元素以外、半金属元素:金属と非金属との境界付近の元素(両方の性質)
遷移元素(横の類似性):3~12族、3族のうち原子番号=57~70と原子番号=89~102は内部遷移元素
典型元素(縦の類似性):遷移元素以外、典型元素には非金属元素と金属元素がある、テネシンも典型元素に含む
内部遷移元素:原子番号=57~70:ランタノイド(La~Yb)、原子番号=89~102:アクチノイド(Ac~No)
希土類元素(17元素):Scスカンジウム,Yイットリウム,ランタノイド=(「ランタン」+「もどき」)
貴ガスは量的に少ないので希ガスとも言われる化合反応を起こしにくい不活性ガス(最外殻までの電子が満杯状態)
2005年に英語表記がrare gasからnoble gasに改められ,日本語での表記も希ガスから貴ガスへと変更になりました
令和3年度の教科書より、日本化学会の提言も踏まえて高貴なガスという意味から貴ガスと表記を変更しています
化学反応しやすい:電子が満杯+1個余計(Na+)、電子が満杯-1個少ない(Cl-)=化学反応を起こしてNaClになる
第12族から第16族は先頭元素名で呼ばれる(亜鉛族,ホウ素族,炭素族,窒素族,酸素族)
ニクトゲン:15族に位置する5つの元素(N,P,As,Sb,Bi),カルコゲン:16族に位置する5つの元素(O,S,Se,Te,Po)
人工元素:天然に存在する元素は原則、原子番号92Uウランまでですが、原子番号92までのうち4つの例外元素
=43Tcテクネチウム,61Pmプロメチウム,85Atアスタチン,87Frフランシウム(同位体)および原子番号が93以上の
超ウラン元素(アクチノイドの一部93~103,104-118の24個),いずれも放射性元素
|
殻 ---spdf軌道---
K 1s L 2s 2p M 3s 3p 3d N 4s 4p 4d 4f O 5s 5p 5d 5f P 6s 6p 6d 6f Q 7s 7p 7d 7f 2 6 10 14 |
電子殻は内側から、K殻、L殻、M殻、N殻…です 各電子殻に入ることのできる電子の数は限りがあって、 K殻に2個、L殻に8個、M殻に18個、N殻に32個となります 電子が電子殻に入るとき、その入り方には規則性(電子配置)がある 規則性①‥電子は普通内側にある電子殻(K殻)から順に入る 内側から入った電子が電子殻の収容できる数を超えるとその外側の電子殻に入る 規則性②‥最外殻電子の数が8個を超えてはいけない(ただし、Pdは例外) 原子は安定な電子殻が多い方を目指す(Ca20の場合、K2,L8,M10でなく、K2,L8,M8,N2) 遷移元素はこの規則性に従わない(後述の例外1,例外2,例外3) |
殻 K L L M M N M N O N O P N O P Q O P Q P Q Q
軌道1s→2s→2p→3s→3p→4s→3d→4p→5s→4d→5p→6s→4f→5d→6p→7s→5f→6d→7p→6f→7d→7f
(E)1s<2s<2p<3s<3p<4s<3d<4p<5s<4d<5p<6s<4f<5d<6p<7s<5f<6d<7p<6f<7d<7f
という順で詰まっていきます(銅などいくつかの例外もある)
○主量子数:n = 1,2,3,…,7 (正の整数) = 電子殻:K,L,M,N,…殻
○主量子数nと方位量子数l(エル)との関係…0≦l≦n-1
○方位量子数l(エル)と磁気量子数mlとの関係…-l≦ml≦l
エネルギー準位(E)は、通常、主量子数(n)が大きくなるほど高く、主量子数(n)が同じ値の場合には、
方位量子数(l:エル)の値が大きくなるほど高いが、n≧3の場合は、高低の逆転が生じる(4s<3d)
このように見ると、M殻の3d軌道の前にN殻の4s軌道が詰まるなど「K殻から順に」のルールが崩れています
実はこれこそが「遷移元素」が存在する理由です
電子軌道は、エネルギーの低い順に収容される(4s<3d)
例として原子番号21、電子数21のスカンジウムを見ていきます
カルシウムは電子数20ですから、電子1個を残して全てカルシウムと同じように詰まります
カルシウムはK殻に2個、L殻に8個、M殻に8個入ると、あと10個の電子がM殻に入れるのですが、
残り2個の電子は、M殻より軌道エネルギーの低いN殻に先に入ります(4s<3d)
(Ca20の場合、K2,L8,M10でなく、K2,L8,M8,N2)
K L L M M N
20Ca:1s2→2s2→2p6→3s2→3p6→4s2
電子数 2 4 10 12 18 20
| s軌道 | ┌────p軌道────┐ | ┌──────d軌道──────┐ | ┌───f軌道───┐ | ||||||||||||||||||||||||
| 周期1 | K殻 | ↑ | ↓ | ||||||||||||||||||||||||
| 周期2 | L殻 | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ||||||||||||||||||
| 周期3 | M殻 | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ||||||||||||||||||
| 周期4 | N殻 | ↑ | ↓ | … | |||||||||||||||||||||||
| : | |||||||||||||||||||||||||||
| ↑ | ↓ |
| ↑ | ↑ | ↑ |
K L L M M N M
21Sc:1s2→2s2→2p6→3s2→3p6→4s2→3d1
電子数 2 4 10 12 18 20 21
この先最外殻電子は4s軌道の2個のまま、3d軌道に入っていくことになり、これが「遷移元素」が生じる原因です
| s軌道 | ┌────p軌道────┐ | ┌──────d軌道──────┐ | ┌───f軌道───┐ | ||||||||||||||||||||||||
| 周期1 | K殻 | ↑ | ↓ | ||||||||||||||||||||||||
| 周期2 | L殻 | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ||||||||||||||||||
| 周期3 | M殻 | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | |||||||||||||||||
| 周期4 | N殻 | ↑ | ↓ | … | |||||||||||||||||||||||
| : | |||||||||||||||||||||||||||
亜鉛族元素:第12族元素は、d軌道が閉殻状態ゆえ典型元素的性質を持つため、以前は「典型元素」に分類され
ていましたが、IUPACの最近の勧告にあわせて、日本も2022年度からは「遷移元素」に含めるとしています
従来の周期表の中には、(La~Lu)及び((Ac~Lr)の各15元素を「内部遷移元素」としているものもありますが、
電子配置からするとLu及びLrは「内部遷移元素」でなく「遷移元素」です
本来、内部遷移元素は(n-2)f1-14の電子を持ち、各シリーズ14個の元素からなることが特長です
ランタノイド(La~Lu)15個のうちLuを除外した(La~Yb)が内部遷移元素14個、Luは遷移元素
(La~Yb)+(Lu~Hg):(14+10)元素=4f+5d
アクチノイド(Ac~Lr)15個のうちLrを除外した(Ac~No)が内部遷移元素14個、Lrは遷移元素
(Ac~No)+(Lr~Cn):(14+10)元素=5f+6d
1番外側の電子殻に収容されている電子のことを最外殻電子といいます
貴ガスの最外殻電子はヘリウムだけ2個で、他の貴ガスはすべて最外殻電子は8個で安定な電子配置をしています
このように最大数の電子が収容された電子殻を閉殻といいます
化学結合や化学的性質に関与する電子のことを価電子といいます
最外殻電子がちょうどその電子殻の最大収容数の場合、または最外殻電子が8個の場合、価電子の数は0とする
貴ガスの場合は最外殻までの電子が満杯状態で、化学反応をそもそも起こさないので、価電子の数はゼロです
価電子の定義は曖昧です(貴ガスは反応性が低いから価電子数はすべて0としますと言う程度の定義です!)
同一元素の原子はその原子核に必ず同数の陽子を持っているが、中性子数の異なる原子核(核種)を同位体と呼ぶ
同位体は陽子と軌道電子の数が等しいため化学的性質はほぼ等しいが、物理的性質は異なる
同位体のうち放射性を有するものを放射性同位体、放射性を有しないものを安定同位体と呼ぶ
| 族→ 周期 |
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 |
| 1 | 1 水素 1766 | 2 ヘリウム 1868 | ||||||||||||||||
| 2 | 3 リチウム 1817 |
4 ベリリウム 1798 |
上段:原子番号、中段:日本名、下段:発見年 | 5 ホウ素 1808 |
6 炭素 古代 |
7 窒素 1772 |
8 酸素 1774 |
9 フッ素 1886 |
10 ネオン 1898 | |||||||||
| 3 | 11 ナトリウム 1807 |
12 マグネシウム 1755 | 13 アルミニウム 1825 |
14 ケイ素 1823 |
15 リン 1669 |
16 硫黄 古代 |
17 塩素 1774 |
18 アルゴン 1894 | ||||||||||
| 4 | 19 カリウム 1807 |
20 カルシウム 1808 |
21 スカンジウム 1879 |
22 チタン 1795 |
23 バナジウム 1801 |
24 クロム 1797 |
25 マンガン 1774 |
26 鉄 古代 |
27 コバルト 1737 |
28 ニッケル 1751 |
29 銅 古代 |
30 亜鉛 古代 |
31 ガリウム 1875 |
32 ゲルマニウム 1886 |
33 ヒ素 1250 |
34 セレン 1817 |
35 臭素 1826 |
36 クリプトン 1898 |
| 5 | 37 ルビジウム 1860 |
38 ストロンチウム 1793 |
39 イットリウム 1794 |
40 ジルコニウム 1789 |
41 ニオブ 1846 |
42 モリブデン 1778 |
43 テクネチウム 1937 |
44 ルテニウム 1844 |
45 ロジウム 1803 |
46 パラジウム 1803 |
47 銀 古代 |
48 カドミウム 1817 |
49 インジウム 1863 |
50 スズ 古代 |
51 アンチモン 1450 |
52 テルル 1782 |
53 ヨウ素 1811 |
54 キセノン 1898 |
| 6 | 55 セシウム 1860 |
56 バリウム 1774 |
ランタ ノイド |
72 ハフニウム 1923 |
73 タンタル 1802 |
74 タングステン 1783 |
75 レニウム 1925 |
76 オスミウム 1803 |
77 イリジウム 1803 |
78 白金 1741 |
79 金 古代 |
80 水銀 古代 |
81 タリウム 1860 |
82 鉛 古代 |
83 ビスマス 1753 |
84 ポロニウム 1898 |
85 アスタチン 1940 |
86 ラドン 1898 |
| 7 | 87 フランシウム 1939 |
88 ラジウム 1898 |
アクチ ノイド |
104 ラザホージウム 1964 |
105 ドブニウム 1970 |
106 シーボーギウム 1974 |
107 ボーリウム 1976 |
108 ハッシウム 1984 |
109 マイトネリウム 1982 |
110 ダームスタチウム 1994 |
111 レントゲニウム 1994 |
112 コペルニシウム 1996 |
113 ニホニウム 2004 |
114 フレロビウム 1999 |
115 モスコビウム 2004 |
116 リバモリウム 2000 |
117 テネシン 2009 |
118 オガネソン 2003 |
| ランタ ノイド |
57 ランタン 1839 |
58 セリウム 1803 |
59 プラセオジム 1885 |
60 ネオジム 1885 |
61 プロメチウム 1945 |
62 サマリウム 1879 |
63 ユウロピウム 1901 |
64 ガドリニウム 1880 |
65 テルビウム 1843 |
66 ジスプロシウム 1886 |
67 ホルミウム 1879 |
68 エルビウム 1843 |
69 ツリウム 1879 |
70 イッテルビウム 1878 |
71 ルテチウム 1907 |
| アクチ ノイド |
89 アクチニウム 1899 |
90 トリウム 1828 |
91 プロトアクチニウム 1917 |
92 ウラン 1789 |
93 ネプツニウム 1940 |
94 プルトニウム 1941 |
95 アメリシウム 1945 |
96 キュリウム 1944 |
97 バークリウム 1949 |
98 カリホルニウム 1950 |
99 アインスタイニウム 1952 |
100 フェルミウム 1953 |
101 メンデレビウム 1955 |
102 ノーベリウム 1958 |
103 ローレンシウム 1961 |
元素数 = 10個、 1個、 1個、 1個、 21個、 49個、 30個、 5個
地表露出:有用資源の採掘
熱分解・電気分解:化合物を単体に分解
分光器:分光分析、スペクトル分析
周期律の発見:三つ組元素、未知元素の予言(周期表の空欄)
放射線:放射性元素の発見
加速器:原子核反応、人工元素、現代の錬金術
| 族→ 周期 |
Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | 0 | ||
| 1 | 1 H 水素 | 2 He ヘリウム | ||||||||
| 2 | 3 Li リチウム |
4 Be ベリリウム |
上段:原子番号(陽子数) 中段:元素記号 |
5 B ホウ素 |
6 C 炭素 |
7 N 窒素 |
8 O 酸素 |
9 F フッ素 |
10 Ne ネオン | |
| 3 | 11 Na ナトリウム |
12 Mg マグネシウム |
(文字色は常温の相) 下段:日本名 |
13 Al アルミニウム |
14 Si ケイ素 |
15 P リン |
16 S 硫黄 |
17 Cl 塩素 |
18 Ar アルゴン | |
| 4 | 19 K カリウム |
20 Ca カルシウム |
21~30 Sc~Zn スカンジウム~亜鉛 |
31 Ga ガリウム |
32 Ge ゲルマニウム |
33 As ヒ素 |
34 Se セレン |
35 Br 臭素 |
36 Kr クリプトン | |
| 5 | 37 Rb ルビジウム |
38 Sr ストロンチウム |
39~48 Y~Cd イットリウム~カドミウム |
49 In インジウム |
50 Sn スズ |
51 Sb アンチモン |
52 Te テルル |
53 I ヨウ素 |
54 Xe キセノン | |
| 6 | 55 Cs セシウム |
56 Ba バリウム |
57~70 La~Yb ランタン~イッテルビウム |
71~80 Lu~Hg ルテチウム~水銀 |
81 Tl タリウム |
82 Pb 鉛 |
83 Bi ビスマス |
84 Po ポロニウム |
85 At アスタチン |
86 Rn ラドン |
| 7 | 87 Fr フランシウム |
88 Ra ラジウム |
89~102 Ac~No アクチニウム~ノーベリウム |
103~112 Lr~Cn ローレンシウム~コペルニシウム |
113 Nh ニホニウム |
114 Fl フレロビウム |
115 Mc モスコビウム |
116 Lv リバモリウム |
117 Ts テネシン |
118 Og オガネソン |
最外殻電子の数が同じ元素は同じ族にまとめた周期表
第4軌道以降は、p軌道の前の隙間(d軌道やf軌道)に電子が入り込み、
元素はどれも同じような性質を示す
メンデレーエフの第二の周期表をベースに貴ガスやLn(57-71)An(89-103)を追加
亜族を追加(A&B)した短周期型周期表
IB,ⅡB,ⅢA–ⅦA,O族をまるごとⅧ族の右に引越した短周期型周期表
亜族問題の解決とブロック(spdf)の分類を同時に達成
族の名称を1–18族と呼ぶことに改めた長周期型周期表
1–18族(縦の同族元素のグループ)
短周期型周期表の改良版
左端のⅠ-Ⅱと右端のⅢ-Oとの間のⅢ-Ⅷ,Ⅰ-Ⅱ族(f軌道とd軌道)の族名を消去
f軌道とd軌道のブロックのグループ化(縦の同族元素のグループの例外)、遷移元素
超長周期型周期表
第4軌道以降は、p軌道の前の隙間(d軌道やf軌道)に電子が入り込み、
元素はどれも同じような性質を示す
メンデレーエフの第二の周期表をベースに貴ガスやLn(57-71)An(89-103)を追加
亜族を追加(A&B)した短周期型周期表
| 族→ 周期 | Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | O | Ⅷ | Ⅷ | Ⅷ | |||||||||||
| A | B | A | B | A | B | A | B | A | B | A | B | A | B | |||||||||
| 1 | H | He | ||||||||||||||||||||
| 2 | Li | Be | B | C | N | O | F | Ne | ||||||||||||||
| 3 | Na | Mg | Al | Si | P | S | Cl | Ar | ||||||||||||||
| 4 | K | Cu | Ca | Zn | Sc | Ga | Ti | Ge | V | As | Cr | Se | Mn | Br | Kr | Fe | Co | Ni | ||||
| 5 | Rb | Ag | Sr | Cd | Y | In | Sn | Nb | Sb | Mo | Te | Tc | I | Xe | Ru | Rh | Pd | |||||
| 6 | Cs | Au | Ba | Hg | La~Lu | Tl | Hf | Pb | Ta | Bi | W | Po | Re | At | Rn | Os | Ir | Pt | ||||
| 7 | Fr | Rg | Ra | Cn | Ac~Lr | Nh | Rf | Fl | Db | Mc | Sg | Lv | Bh | Ts | Og | Hs | Mt | Ds | ||||
亜族問題の解決とブロック(spdf)の分類を同時に達成
| 族→ 周期 | Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | Ⅷ | Ⅷ | Ⅷ | Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | O |
| 1 | H | He | ||||||||||||||||
| 2 | Li | Be | B | C | N | O | F | Ne | ||||||||||
| 3 | Na | Mg | Al | Si | P | S | Cl | Ar | ||||||||||
| 4 | K | Ca | Sc | Ti | Ⅴ | Cr | Mn | Fe | Co | Ni | Cu | Zn | Ga | Ge | As | Se | Br | Kr |
| 5 | Rb | Sr | Y | Zr | Nb | Mo | Tc | Ru | Rh | Pd | Ag | Cd | In | Sn | Sb | Te | I | Xe |
| 6 | Cs | Ba | La~Lu | Hf | Ta | W | Re | Os | Ir | Pt | Au | Hg | Tl | Pb | Bi | Po | At | Rn |
| 7 | Fr | Ra | Ac~Lr | Rf | Db | Sg | Bh | Hs | Mt | Ds | Rg | Cn | Nh | Fl | Mc | Lv | Ts | Og |
1–18族(縦の同族元素のグループ)
| 価電子数 | 1 | 2 | 1か2 | 3 | 4 | 5 | 6 | 7 | 0 | |||||||||
| 族→ 周期 |
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 |
| 1 | H | He | ||||||||||||||||
| 2 | Li | Be | B | C | N | O | F | Ne | ||||||||||
| 3 | Na | Mg | Al | Si | P | S | Cl | Ar | ||||||||||
| 4 | K | Ca | Sc | Ti | Ⅴ | Cr | Mn | Fe | Co | Ni | Cu | Zn | Ga | Ge | As | Se | Br | Kr |
| 5 | Rb | Sr | Y | Zr | Nb | Mo | Tc | Ru | Rh | Pd | Ag | Cd | In | Sn | Sb | Te | I | Xe |
| 6 | Cs | Ba | La~Lu | Hf | Ta | W | Re | Os | Ir | Pt | Au | Hg | Tl | Pb | Bi | Po | At | Rn |
| 7 | Fr | Ra | Ac~Lr | Rf | Db | Sg | Bh | Hs | Mt | Ds | Rg | Cn | Nh | Fl | Mc | Lv | Ts | Og |
左端のⅠ-Ⅱと右端のⅢ-Oとの間のⅢ-Ⅷ,Ⅰ-Ⅱ族(f軌道とd軌道)の族名を消去
f軌道とd軌道のブロックのグループ化(縦の同族元素のグループの例外)、遷移元素
| 族→ 周期 |
Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | 0 | ||
| 1 | 2 | 3 | 3-12 | 13 | 14 | 15 | 16 | 17 | 18 | |
| 1 | H | He | ||||||||
| 2 | Li | Be | B | C | N | O | F | Ne | ||
| 3 | Na | Mg | Al | Si | P | S | Cl | Ar | ||
| 4 | K | Ca | Sc~Zn | Ga | Ge | As | Se | Br | Kr | |
| 5 | Rb | Sr | Y~Cd | In | Sn | Sb | Te | I | Xe | |
| 6 | Cs | Ba | La~Yb | Lu~Hg | Tl | Pb | Bi | Po | At | Rn |
| 7 | Fr | Ra | Ac~No | Lr~Cn | Nh | Fl | Mc | Lv | Ts | Og |
| A(2) | 内部遷移元素C(14) | 遷移元素B(10) | 典型元素A(6) | |||||||||||||||||||||||||||||
| 族→ 周期 | ⅠA | ⅡA | ⅠC~ⅩⅣC | ⅠB~ⅩB | ⅢA~ⅧA | |||||||||||||||||||||||||||
| 1 | 2 | 3 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | ||||||||||||||
| 1 | H | He | ||||||||||||||||||||||||||||||
| 2 | Li | Be | B | C | N | O | F | Ne | ||||||||||||||||||||||||
| 3 | Na | Mg | Al | Si | P | S | Cl | Ar | ||||||||||||||||||||||||
| 4 | K | Ca | Sc | Ti | Ⅴ | Cr | Mn | Fe | Co | Ni | Cu | Zn | Ga | Ge | As | Se | Br | Kr | ||||||||||||||
| 5 | Rb | Sr | Y | Zr | Nb | Mo | Tc | Ru | Rh | Pd | Ag | Cd | In | Sn | Sb | Te | I | Xe | ||||||||||||||
| 6 | Cs | Ba | La | Ce | Pr | Nd | Pm | Sm | Eu | Gd | Tb | Dy | Ho | Er | Tm | Yb | Lu | Hf | Ta | W | Re | Os | Ir | Pt | Au | Hg | Tl | Pb | Bi | Po | At | Rn |
| 7 | Fr | Ra | Ac | Th | Pa | U | Np | Pu | Am | Cm | Bk | Cf | Es | Fm | Md | No | Lr | Rf | Db | Sg | Bh | Hs | Mt | Ds | Rg | Cn | Nh | Fl | Mc | Lv | Ts | Og |
|
左記の表から特定の元素の電子数と
原子番号が求められます 周期 元素数 軌道 1 2 1s 2 8 2s 2p 3 8 3s 3p 4 18 4s 3d 4p 5 18 5s 4d 5p 6 32 6s 4f 5d 6p 7 32 7s 5f 6d 7p abc=電子殻a,電子軌道bの電子数がc個 (a:1~7,b:spdf,c:1~14) 電子殻a=1~7:K~Q、 電子軌道bの軌道数=s:1個,p:3個,d:5個,f:7個 軌道電子数最大 1x2 ,3x2 ,5x2 ,7x2 軌道電子数最大 s:2 ,p:6 ,d:10 ,f:14 例:34Seセレン 周期1:1s2 周期2:2s2,2p6 周期3:3s2,3p6 周期4:4s2,3d10,4p4 cの合計=1s2,2s2,2p6,3s2,3p6,4s2,3d10,4p4 =34 よって、セレンの電子数=34:原子番号 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TOP|貴ガスの電子配置|TOP | |||||||
|
◆引用の明示:Wikipedia◆
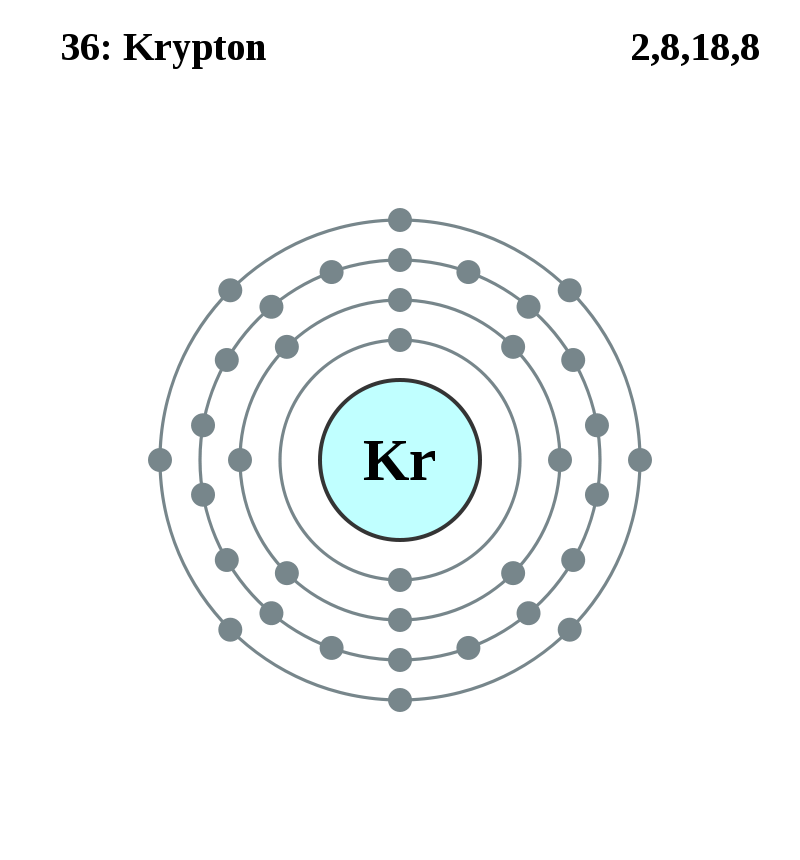
2=2 2+8=10 2+8+8=18 2+8+18+8=36
| |||||||
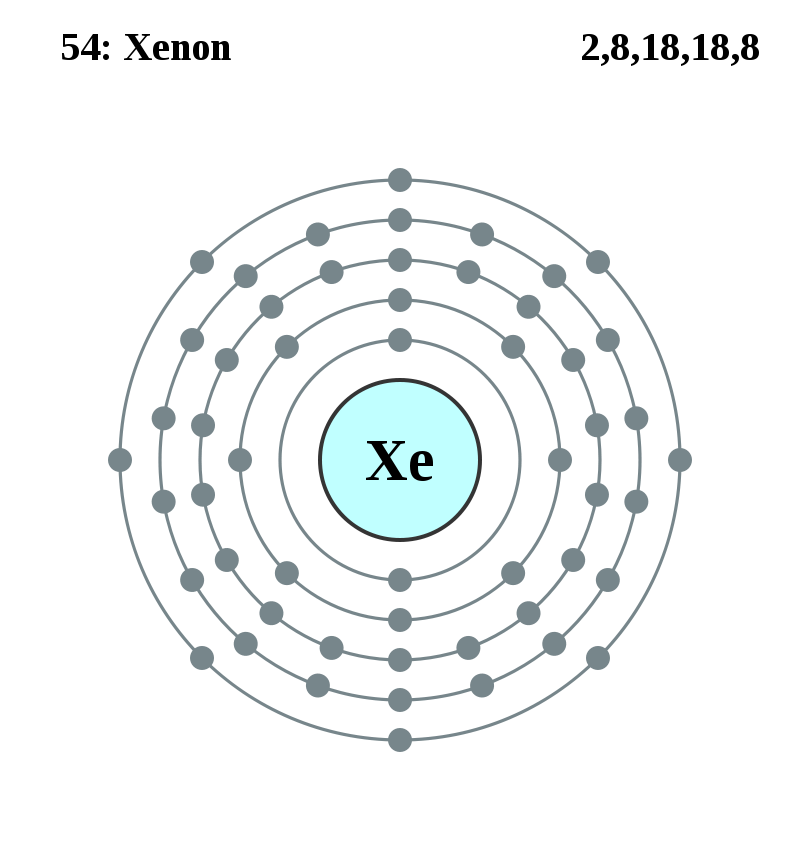
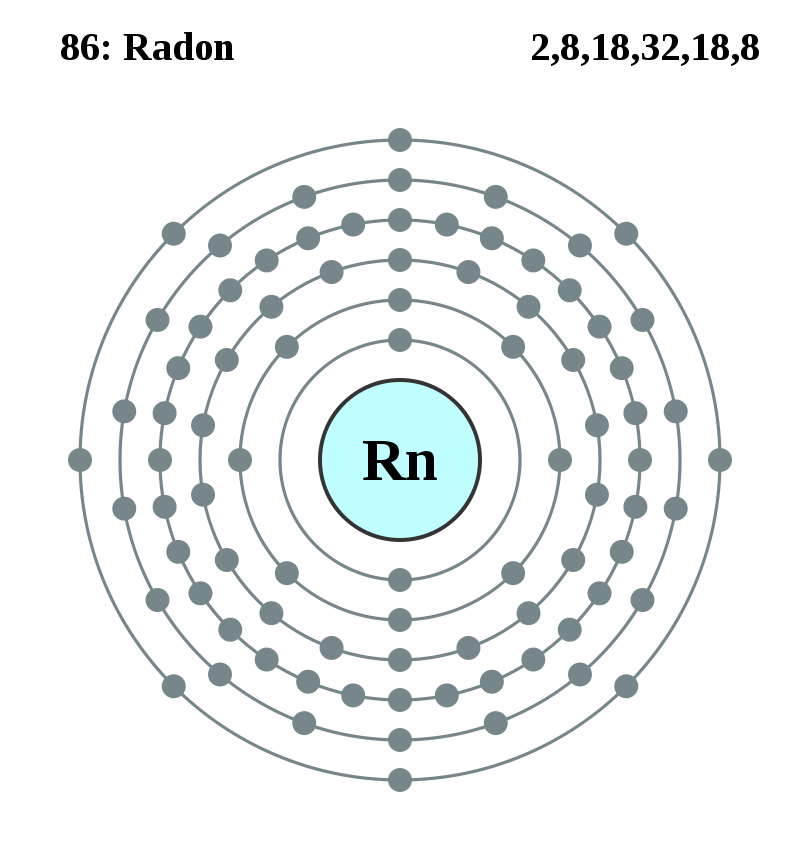
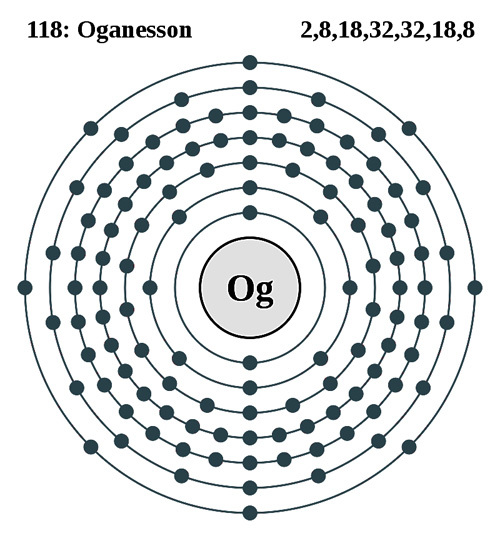
2+8+18+18+8=54 2+8+18+32+18+8=86 2+8+18+32+32+18+8=118
| |||||||
|
電子が軌道に入るときは、次の規則がある ①構成原理 :エネルギーの低い軌道から順に入っていく (例外あり) ②パウリの排他律:1つの軌道には電子が2個まで入ることができる(スピン= +1/2,-1/2) ③フントの規則 :複数の軌道があるときは電子はできるだけ別々の軌道にスピンの向きを揃えて入っていく 電子軌道bの軌道数=s:1個,p:3個,d:5個,f:7個 軌道電子数最大 1x2 ,3x2 ,5x2 ,7x2 | |||||||
|
最外殻電子数:1個~8個 最外殻電子数が8個の場合が貴ガス・不活性ガス 以下は、原子番号118のOgオガネソンの電子配置です ← [周期表の仕組み]の表のab行から後記(1) p軌道の前の隙間d軌道やf軌道に電子が入り込み:低いエネルギ―軌道 1s2,2s2,2p6,3s2,3p6,4s2,3d10,4p6,5s2,4d10,5p6,6s2,4f14,5d10,6p6,7s2,5f14,6d10,7p6 … (1) 1 2 3 4 5 6 7 ← 周期1~7 abc=電子殻a,電子軌道bの電子数がc個(a:1~7,b:spdf,c:1~14) 電子殻a=1~7:K~Q、軌道電子数最大=s:2,p:6,d:10,f:14、前記(1)を電子殻aで並べ直すと次行 1s2,2s2,2p6,3s2,3p6,3d10,4s2,4p6,4d10,4f14,5s2,5p6,5d10,5f14,6s2,6p6,6d10,7s2,7p6 K L M N O P Q ← 電子殻K~Q オガネソン=原子核に近い内側からK殻~Q殻=軌道殻7 電子配置 :K殻,L殻,M殻,N殻,O殻,P殻,Q殻 主量子数 : 1, 2, 3, 4, 5, 6, 7 電子収容数: 2, 8, 18, 32, 32, 18, 8 Σc 軌道の数 : 1, 4, 9, 16, 16, 9, 4 電子収容数/2 ●電子の数=2+8+18+32+32+18+8=118:オガネソンの原子番号 電子殻K~Q=K L M N O P Q ●元素の数=2+8+8+18+18+32+32=118:オガネソンの原子番号 周期1~7=1 2 3 4 5 6 7 | |||||||
|
ヘリウム=2 2 1s2
ネオン=2,8 10 1s2,2s2,2p6 アルゴン=2,8,8 18 1s2,2s2,2p6,3s2,3p6 クリプトン=2,8,18,8 36 1s2,2s2,2p6,3s2,3p6,3d10,4s2,4p6 キセノン=2,8,18,18,8 54 1s2,2s2,2p6,3s2,3p6,3d10,4s2,4p6,4d10 ,5s2,5p6 ラドン=2,8,18,32,18,8 86 1s2,2s2,2p6,3s2,3p6,3d10,4s2,4p6,4d10,4f14,5s2,5p6,5d10 ,6s2,6p6 オガネソン=2,8,18,32,32,18,8 118 1s2,2s2,2p6,3s2,3p6,3d10,4s2,4p6,4d10,4f14,5s2,5p6,5d10,5f14,6s2,6p6,6d10,7s2,7p6 電子殻K~Q=K L M N O P Q K L M N O P Q | |||||||
|
ヘリウム=2 2 1s2
ネオン=2,8 10 1s2,2s2,2p6 アルゴン=2,8,8 18 1s2,2s2,2p6,3s2,3p6 クリプトン=2,8,8,18 36 1s2,2s2,2p6,3s2,3p6,4s2,3d10,4p6 キセノン=2,8,8,18,18 54 1s2,2s2,2p6,3s2,3p6,4s2,3d10,4p6,5s2,4d10,5p6 ラドン=2,8,8,18,18,32 86 1s2,2s2,2p6,3s2,3p6,4s2,3d10,4p6,5s2,4d10,5p6,6s2,4f14,5d10,6p6 オガネソン=2,8,8,18,18,32,32 118 1s2,2s2,2p6,3s2,3p6,4s2,3d10,4p6,5s2,4d10,5p6,6s2,4f14,5d10,6p6,7s2,5f14,6d10,7p6 周期1~7=1 2 3 4 5 6 7 1 2 3 4 5 6 7 |
|||||||
| 最大電子数(殻) | 2 | 8 | 18 | 32 | 32 | 18 | 8 | |||||||||||||||
| 最大電子数(副殻) | 2 | 2 | 6 | 2 | 6 | 10 | 2 | 6 | 10 | 14 | 2 | 6 | 10 | 14 | 2 | 6 | 10 | 2 | 6 | |||
| 族 | 元素記号 | 元素名 | 原子 番号 | 1s | 2s | 2p | 3s | 3p | 3d | 4s | 4p | 4d | 4f | 5s | 5p | 5d | 5f | 6s | 6p | 6d | 7s | 7p |
| K | L | M | N | O | P | Q | ||||||||||||||||
| 第1周期 | 1s | 2s | 2p | 3s | 3p | 3d | 4s | 4p | 4d | 4f | 5s | 5p | 5d | 5f | 6s | 6p | 6d | 7s | 7p | |||
| 1 | H | 1 | 1 | |||||||||||||||||||
| 18 | He | 2 | 2 | |||||||||||||||||||
| 第2周期 | 1s | 2s | 2p | 3s | 3p | 3d | 4s | 4p | 4d | 4f | 5s | 5p | 5d | 5f | 6s | 6p | 6d | 7s | 7p | |||
| 1 | Li | 3 | 2 | 1 | ||||||||||||||||||
| 2 | Be | 4 | 2 | 2 | ||||||||||||||||||
| 13 | B | 5 | 2 | 2 | 1 | |||||||||||||||||
| 14 | C | 6 | 2 | 2 | 2 | |||||||||||||||||
| 15 | N | 7 | 2 | 2 | 3 | |||||||||||||||||
| 16 | O | 8 | 2 | 2 | 4 | |||||||||||||||||
| 17 | F | 9 | 2 | 2 | 5 | |||||||||||||||||
| 18 | Ne | 10 | 2 | 2 | 6 | |||||||||||||||||
| 第3周期 | 1s | 2s | 2p | 3s | 3p | 3d | 4s | 4p | 4d | 4f | 5s | 5p | 5d | 5f | 6s | 6p | 6d | 7s | 7p | |||
| 1 | Na | 11 | 10 1s … 2p ネオン核 [Ne] | 1 | ||||||||||||||||||
| 2 | Mg | 12 | 2 | |||||||||||||||||||
| 13 | Al | 13 | 2 | 1 | ||||||||||||||||||
| 14 | Si | 14 | 2 | 2 | ||||||||||||||||||
| 15 | P | 15 | 2 | 3 | ||||||||||||||||||
| 16 | S | 16 | 2 | 4 | ||||||||||||||||||
| 17 | Cl | 17 | 2 | 5 | ||||||||||||||||||
| 18 | Ar | 18 | 2 | 6 | ||||||||||||||||||
| 第4周期 | 1s | 2s | 2p | 3s | 3p | 3d | 4s | 4p | 4d | 4f | 5s | 5p | 5d | 5f | 6s | 6p | 6d | 7s | 7p | |||
| 1 | K | 19 | 18 1s … 3p アルゴン核 [Ar] | 1 | ||||||||||||||||||
| 2 | Ca | 20 | 2 | |||||||||||||||||||
| 3 | Sc | 21 | 1 | 2 | ||||||||||||||||||
| 4 | Ti | 22 | 2 | 2 | ||||||||||||||||||
| 5 | V | 23 | 3 | 2 | ||||||||||||||||||
| 6 | Cr | 24 | 5 | 1 | ||||||||||||||||||
| 7 | Mn | 25 | 5 | 2 | ||||||||||||||||||
| 8 | Fe | 26 | 6 | 2 | ||||||||||||||||||
| 9 | Co | 27 | 7 | 2 | ||||||||||||||||||
| 10 | Ni | 28 | 8 | 2 | ||||||||||||||||||
| 11 | Cu | 29 | 10 | 1 | ||||||||||||||||||
| 12 | Zn | 30 | 10 | 2 | ||||||||||||||||||
| 13 | Ga | 31 | 10 | 2 | 1 | |||||||||||||||||
| 14 | Ge | 32 | 10 | 2 | 2 | |||||||||||||||||
| 15 | As | 33 | 10 | 2 | 3 | |||||||||||||||||
| 16 | Se | 34 | 10 | 2 | 4 | |||||||||||||||||
| 17 | Br | 35 | 10 | 2 | 5 | |||||||||||||||||
| 18 | Kr | 36 | 10 | 2 | 6 | |||||||||||||||||
| 第5周期 | 1s | 2s | 2p | 3s | 3p | 3d | 4s | 4p | 4d | 4f | 5s | 5p | 5d | 5f | 6s | 6p | 6d | 7s | 7p | |||
| 1 | Rb | 37 | 36 1s … 4p クリプトン核 [Kr] | 1 | ||||||||||||||||||
| 2 | Sr | 38 | 2 | |||||||||||||||||||
| 3 | Y | 39 | 1 | 2 | ||||||||||||||||||
| 4 | Zr | 40 | 2 | 2 | ||||||||||||||||||
| 5 | Nb | 41 | 4 | 1 | ||||||||||||||||||
| 6 | Mo | 42 | 5 | 1 | ||||||||||||||||||
| 7 | Tc | 43 | 5 | 2 | ||||||||||||||||||
| 8 | Ru | 44 | 7 | 1 | ||||||||||||||||||
| 9 | Rh | 45 | 8 | 1 | ||||||||||||||||||
| 10 | Pd | 46 | 10 | |||||||||||||||||||
| 11 | Ag | 47 | 10 | 1 | ||||||||||||||||||
| 12 | Cd | 48 | 10 | 2 | ||||||||||||||||||
| 13 | In | 49 | 10 | 2 | 1 | |||||||||||||||||
| 14 | Sn | 50 | 10 | 2 | 2 | |||||||||||||||||
| 15 | Sb | 51 | 10 | 2 | 3 | |||||||||||||||||
| 16 | Te | 52 | 10 | 2 | 4 | |||||||||||||||||
| 17 | I | 53 | 10 | 2 | 5 | |||||||||||||||||
| 18 | Xe | 54 | 10 | 2 | 6 | |||||||||||||||||
| 第6周期 | 1s | 2s | 2p | 3s | 3p | 3d | 4s | 4p | 4d | 4f | 5s | 5p | 5d | 5f | 6s | 6p | 6d | 7s | 7p | |||
| 1 | Cs | 55 | 54=68-14 1s … 5p-(4f) キセノン核 [Xe] | 1 | ||||||||||||||||||
| 2 | Ba | 56 | 2 | |||||||||||||||||||
| 3 | La | 57 | 1 | 2 | ||||||||||||||||||
| 3 | Ce | 58 | 46=68-(14+2+6) 1s … 5p-(4f,5s,5p) キセノン核 [Xe] | 1 | 2 | 6 | 1 | 2 | ||||||||||||||
| 3 | Pr | 59 | 3 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Nd | 60 | 4 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Pm | 61 | 5 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Sm | 62 | 6 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Eu | 63 | 7 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Gd | 64 | 7 | 2 | 6 | 1 | 2 | |||||||||||||||
| 3 | Tb | 65 | 9 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Dy | 66 | 10 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Ho | 67 | 11 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Er | 68 | 12 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Tm | 69 | 13 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Yb | 70 | 14 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Lu | 71 | 68 1s … 5p キセノン核 [Xe] | 1 | 2 | |||||||||||||||||
| 4 | Hf | 72 | 2 | 2 | ||||||||||||||||||
| 5 | Ta | 73 | 3 | 2 | ||||||||||||||||||
| 6 | W | 74 | 4 | 2 | ||||||||||||||||||
| 7 | Re | 75 | 5 | 2 | ||||||||||||||||||
| 8 | Os | 76 | 6 | 2 | ||||||||||||||||||
| 9 | Ir | 77 | 7 | 2 | ||||||||||||||||||
| 10 | Pt | 78 | 9 | 1 | ||||||||||||||||||
| 11 | Au | 79 | 10 | 1 | ||||||||||||||||||
| 12 | Hg | 80 | 10 | 2 | ||||||||||||||||||
| 13 | Tl | 81 | 10 | 2 | 1 | |||||||||||||||||
| 14 | Pb | 82 | 10 | 2 | 2 | |||||||||||||||||
| 15 | Bi | 83 | 10 | 2 | 3 | |||||||||||||||||
| 16 | Po | 84 | 10 | 2 | 4 | |||||||||||||||||
| 17 | At | 85 | 10 | 2 | 5 | |||||||||||||||||
| 18 | Rn | 86 | 10 | 2 | 6 | |||||||||||||||||
| 第7周期 | 1s | 2s | 2p | 3s | 3p | 3d | 4s | 4p | 4d | 4f | 5s | 5p | 5d | 5f | 6s | 6p | 6d | 7s | 7p | |||
| 1 | Fr | 87 | 78 1s … 5d ラドン核 [Rn] | 2 | 6 | 1 | ||||||||||||||||
| 2 | Ra | 88 | 2 | 6 | 2 | |||||||||||||||||
| 3 | Ac | 89 | 2 | 6 | 1 | 2 | ||||||||||||||||
| 3 | Th | 90 | 2 | 6 | 2 | 2 | ||||||||||||||||
| 3 | Pa | 91 | 2 | 2 | 6 | 1 | 2 | |||||||||||||||
| 3 | U | 92 | 3 | 2 | 6 | 1 | 2 | |||||||||||||||
| 3 | Np | 93 | 4 | 2 | 6 | 1 | 2 | |||||||||||||||
| 3 | Pu | 94 | 6 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Am | 95 | 7 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Cm | 96 | 7 | 2 | 6 | 1 | 2 | |||||||||||||||
| 3 | Bk | 97 | 9 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Cf | 98 | 10 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Es | 99 | 11 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Fm | 100 | 12 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Md | 101 | 13 | 2 | 6 | 2 | ||||||||||||||||
| 3 | No | 102 | 14 | 2 | 6 | 2 | ||||||||||||||||
| 3 | Lr | 103 | 14 | 2 | 6 | 2 | 1 | |||||||||||||||
| 4 | Rf (推定) | 104 | 14 | 2 | 6 | 2 | 2 | |||||||||||||||
| 5 | Db (推定) | 105 | 14 | 2 | 6 | 3 | 2 | |||||||||||||||
| 6 | Sg (推定) | 106 | 14 | 2 | 6 | 4 | 2 | |||||||||||||||
| 7 | Bh (計算値) | 107 | 14 | 2 | 6 | 5 | 2 | |||||||||||||||
| 8 | Hs (推定) | 108 | 14 | 2 | 6 | 6 | 2 | |||||||||||||||
| 9 | Mt | 109 | 14 | 2 | 6 | 7 | 2 | |||||||||||||||
| 10 | Ds | 110 | 14 | 2 | 6 | 9 | 1 | |||||||||||||||
| 11 | Rg | 111 | 14 | 2 | 6 | 9 | 2 | |||||||||||||||
| 12 | Cn | 112 | 14 | 2 | 6 | 10 | 2 | |||||||||||||||
| 13 | Nh | 113 | 14 | 2 | 6 | 10 | 2 | 1 | ||||||||||||||
| 14 | Fl (推定) | 114 | 14 | 2 | 6 | 10 | 2 | 2 | ||||||||||||||
| 15 | Mc (推定) | 115 | 14 | 2 | 6 | 10 | 2 | 3 | ||||||||||||||
| 16 | Lv (推定) | 116 | 14 | 2 | 6 | 10 | 2 | 4 | ||||||||||||||
| 17 | Ts (計算値) | 117 | 14 | 2 | 6 | 10 | 2 | 5 | ||||||||||||||
| 18 | Og (推定) | 118 | 14 | 2 | 6 | 10 | 2 | 6 | ||||||||||||||
| 族 | 元素記号 | 元素名 | 原子 番号 | 1s | 2s | 2p | 3s | 3p | 3d | 4s | 4p | 4d | 4f | 5s | 5p | 5d | 5f | 6s | 6p | 6d | 7s | 7p |
| K | L | M | N | O | P | Q | ||||||||||||||||
| 最大電子数(副殻) | 2 | 2 | 6 | 2 | 6 | 10 | 2 | 6 | 10 | 14 | 2 | 6 | 10 | 14 | 2 | 6 | 10 | 2 | 6 | |||
| 最大電子数(殻) | 2 | 8 | 18 | 32 | 32 | 18 | 8 | |||||||||||||||
背景の色=非金属、アルカリ金属、アルカリ土類金属、遷移金属、ハロゲン、貴ガス、典型金属
遷移元素:3~12族、3族のうち原子番号=57~70と原子番号=89~102は内部遷移元素
[貴ガスの元素記号]:電子配置の共通項を簡略形式で表現、例:Se=[Ar],3d10,4s2,4p4
クロム(Cr)と銅(Cu)の電子配置は次の通りです
最外殻電子4sから電子を一つ奪って3d軌道に入っています
●クロム(Cr,24):1s2,2s2,2p6,3s2,3p6,3d5,4s1
d軌道に5個の不対電子が入ることで、スピンが同じ方向を向きエネルギー的に安定化します
(d軌道が半閉殻)
| s軌道 | ┌────p軌道────┐ | ┌──────d軌道──────┐ | ┌───f軌道───┐ | ||||||||||||||||||||||||
| 周期1 | K殻 | ↑ | ↓ | ||||||||||||||||||||||||
| 周期2 | L殻 | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ||||||||||||||||||
| 周期3 | M殻 | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↑ | ↑ | ↑ | ↑ | |||||||||||||
| 周期4 | N殻 | ↑ | … | ||||||||||||||||||||||||
d軌道に10個の電子対が入ることで、d軌道の電子が全て互いに逆向きスピンで充填され、
エネルギー的に安定化します(d軌道が閉殻)
| s軌道 | ┌────p軌道────┐ | ┌──────d軌道──────┐ | ┌───f軌道───┐ | ||||||||||||||||||||||||
| 周期1 | K殻 | ↑ | ↓ | ||||||||||||||||||||||||
| 周期2 | L殻 | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ||||||||||||||||||
| 周期3 | M殻 | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ||||||||
| 周期4 | N殻 | ↑ | … | ||||||||||||||||||||||||
最外殻電子5sから電子を二つ奪って4d軌道に入っています
●パラジウム(Pd,46):1s2,2s2,2p6,3s2,3p6,3d10,4s2,4p6,4d10,5s0
先に5s軌道から電子が入り始めますが、4d軌道に電子が入り始め4d軌道の電子が増え、
Pdでは5s(↑↓)4d(↑↓)(↑↓)(↑↓)(↑)(↑)ではなく
| s軌道 | ┌────p軌道────┐ | ┌──────d軌道──────┐ | ┌───f軌道───┐ | ||||||||||||||||||||||||
| 周期1 | K殻 | ↑ | ↓ | ||||||||||||||||||||||||
| 周期2 | L殻 | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ||||||||||||||||||
| 周期3 | M殻 | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ||||||||
| 周期4 | N殻 | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↑ | … | |||||||||
| 周期5 | О殻 | ↑ | ↓ | … | |||||||||||||||||||||||
| s軌道 | ┌────p軌道────┐ | ┌──────d軌道──────┐ | ┌───f軌道───┐ | ||||||||||||||||||||||||
| 周期1 | K殻 | ↑ | ↓ | ||||||||||||||||||||||||
| 周期2 | L殻 | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ||||||||||||||||||
| 周期3 | M殻 | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ||||||||
| 周期4 | N殻 | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ↓ | … | |||||||
| 周期5 | О殻 | … | |||||||||||||||||||||||||
電子はスピンの向きを揃えたがる性質と、できるだけ離れた電子軌道に入ろうとする二つの重要な性質
があり、最終的に原子全体としていかにバランスよく電子配置するかという観点で電子を配置します
dブロック(3d~6d)の例外1:
d軌道の副殻は、s軌道の副殻から1個の電子(パラジウムの場合は2個)を「借ります」
fブロック(4f~5f)の例外2:
d軌道の副殻は、f軌道の副殻から1個の電子(トリウムの場合は2個)を「借りる」ことがよくあります
pブロック(2p~7p)の例外3:
1970年、ローレンシウム(Lr,103)の基底状態の電子配置は、構造原理に従って、[Rn]5f146d17s2であり、
同族体であるルテチウムの[Xe]4f145d16s2とも合致すると予測された
しかし翌年、この予測に疑義を唱え、その代わり、[Rn]5f147s27p1という異常な電子配置を取るとする
計算結果が公表された
電子の(spdf)軌道とスピン(↑↓)
| 年代 | 物理学者 | アイデア | ||
|---|---|---|---|---|
| 量子論 の誕生 | 1690 | ホイヘンス | 光の波動説 | |
| 17世紀末 | ニュートン | 光の粒子説 | ||
| 19世紀初頭 | ヤング | ヤングの実験 | ||
| 1873 | マクスウェル | 光は電磁波の証明 | ||
| 1888 | ヘルツ | 光電効果を発見 | ||
| 前期量子 論の時代 | 第1期 | 1900年代 | プランク | エネルギー量子仮説 |
| アインシュタイン | 光量子仮説 | |||
| 第2期 | 1910年代 | ラザフォード | 原子核の発見 | |
| ボーア | 原子構造の量子論 | |||
| 現代的な 量子力学 | 第3期 | 1920年代 | ド・ブロイ | 物質波の提唱 |
| ハイゼンベルク | 行列力学 | |||
| シュレディンガー | シュレディンガー方程式 | |||
| ボルン | 物質波は確率の波 | |||
| ハイゼンベルク | 不確定性原理 | |||
| ボーアとハイゼンベルク | コペンハーゲン解釈 | |||
| ディラック | ディラック方程式 | |||
| 量子力学 の発展 | 1930年代~ | エヴェレット | 多世界解釈 | |
| 南部陽一郎 | 対称性の自発的な破れ | |||
| ゲルマンとツヴァイク | クォークモデルを発表 | |||
| ピーター・ヒッグス | ヒッグス機構 | |||
| ワインバーグら | 電弱統一理論 | |||
| ワインバーグと小林・益川 | 標準模型(標準理論) | |||
| ジョージとグラショウ | 大統一理論 | |||
量子論は何人もの天才物理学者がアイデアを出し合って作られた
ホイヘンスの波動説:実体が移動するのでなく、それぞれの地点で振動が起こっている
ニュートンの粒子説:「粒子」という実体がAからBへ移動する
ヤングの二重スリット実験 :光の干渉は波の性質の証拠
マクスウェルの電磁波の予言:光は電磁波の一種である、ヘルツによる電磁波の発見
アインシュタインの光量子説:光電効果は光が粒子なら説明がつく、コンプトン効果
ド・ブロイの物質波 :波と粒子の二面性は光だけでなくほかの物質にもある
コペンハーゲン解釈 :量子は波と粒子の二面性をもつ、量子の相補性
ニュートンの粒子説:「粒子」という実体がAからBへ移動する
ヤングの二重スリット実験 :光の干渉は波の性質の証拠
マクスウェルの電磁波の予言:光は電磁波の一種である、ヘルツによる電磁波の発見
アインシュタインの光量子説:光電効果は光が粒子なら説明がつく、コンプトン効果
ド・ブロイの物質波 :波と粒子の二面性は光だけでなくほかの物質にもある
コペンハーゲン解釈 :量子は波と粒子の二面性をもつ、量子の相補性
「粒子の運動」 ⇒ 「波の塊の移動」
電磁波の「波動性」:波長が長くなるほど、波の回折現象や干渉現象など「波動性」が顕著に表れる
電磁波の「粒子性」:波長が短くなるほど、波の性質が目立たず直進する「粒子性」が顕著に表れる
「粒子性」とは電磁波が数えられることを指し、1個1個のエネルギーの“かたまり”を光子と呼ぶ
光子の運動量p=mc(質量×光速),プランク定数h,波長λ=h/p
光子の質量は0なので質量とエネルギーの等価式E=mc2を用いてm=E/c2,p=E/c,
光子の振動数をνとすると、E=hν,c=νλの関係からp=h/λと変形できます
プランク定数h=6.62607015×10−34Jsは非常に小さいので極微で意味がある
コペンハーゲン解釈:電子の波は電子の発見確率,振幅が大→発見確率が大,振幅が小→発見確率が小
量子の世界の「波と粒子の二重性」は、経験的理解を超え、直観的理解が難しい部分がある
素粒子のスピンの性格、パウリの排他律、トンネル効果、量子もつれ等も同様に難解である
波の性質である波長と粒子の性質であるエネルギーは密接な関係がある
ミクロの世界のエネルギー単位:1eV=1電子を1ボルトで加速時のエネルギー
光子のエネルギーは波長に逆比例する … E=(hc)/λ
3evの紫色の光の波長は0.4ミクロン、2evの赤色の光の波長は0.7ミクロン
電磁波の「粒子性」:波長が短くなるほど、波の性質が目立たず直進する「粒子性」が顕著に表れる
「粒子性」とは電磁波が数えられることを指し、1個1個のエネルギーの“かたまり”を光子と呼ぶ
「粒子性」:位置は精度よく定まるが、波長は不確かになる(波長が短い塊 ⇒ )
「波動性」:波長は精度よく定まるが、位置は不確かになる(波長が長い波 )
光子の実体は、短い(多くの場合数cm~数mの)切れ端のような電磁波(切れ端1個はある波長で振動)「波動性」:波長は精度よく定まるが、位置は不確かになる(波長が長い波 )
光子の運動量p=mc(質量×光速),プランク定数h,波長λ=h/p
光子の質量は0なので質量とエネルギーの等価式E=mc2を用いてm=E/c2,p=E/c,
光子の振動数をνとすると、E=hν,c=νλの関係からp=h/λと変形できます
プランク定数h=6.62607015×10−34Jsは非常に小さいので極微で意味がある
コペンハーゲン解釈:電子の波は電子の発見確率,振幅が大→発見確率が大,振幅が小→発見確率が小
量子の世界の「波と粒子の二重性」は、経験的理解を超え、直観的理解が難しい部分がある
素粒子のスピンの性格、パウリの排他律、トンネル効果、量子もつれ等も同様に難解である
波の性質である波長と粒子の性質であるエネルギーは密接な関係がある
ミクロの世界のエネルギー単位:1eV=1電子を1ボルトで加速時のエネルギー
光子のエネルギーは波長に逆比例する … E=(hc)/λ
3evの紫色の光の波長は0.4ミクロン、2evの赤色の光の波長は0.7ミクロン
特殊相対性理論において、エネルギーEと運動量pの間に、
E=cpなる関係が成立しますので、 … (B)
A式とB式が等価になるためにはA式の質量mは0でなければならないのです
E=mc2という式は物質が消滅(質量=0)すると発生するエネルギーの式で、
アインシュタインがm=E/c2を一般向けに平易に書き換えた式です
E2
――=m2c2+p2なる関係が成立します … (A)
c2
光子の場合、p=mc=E/cからエネルギーEと運動量pの間に――=m2c2+p2なる関係が成立します … (A)
c2
E=cpなる関係が成立しますので、 … (B)
A式とB式が等価になるためにはA式の質量mは0でなければならないのです
E=mc2という式は物質が消滅(質量=0)すると発生するエネルギーの式で、
アインシュタインがm=E/c2を一般向けに平易に書き換えた式です
質量とエネルギーの等価原理
1905年特殊相対性理論
運動を見る視点の側の限定条件:等速運動または静止
光速不変:光の速度はどのような立場から見ても一定である、光よりも速い速度はあり得ない
同時性の破れ:異なる位置における同時性は、慣性系によって変わる(光速不変の原理による)
電車の中央から両壁に光を発射、電車内にいる観測者は両壁到達が同時
外のホームにいる観測者には準光速走行電車の両壁到達は同時ではない
時間の相対性:光の速度に近づくと、時間の流れはゆっくりになる
E=mc2(E:エネルギー、m:質量、c:光速)
質量をエネルギーに、エネルギーを質量に変えられる
1916年一般相対性理論
空間の相対性:重力の大きいところでは空間がゆがむ、大きな重力は光さえも曲げる
時間の相対性:重力の大きいところでは時間の流れはゆっくりになる
運動を見る視点の側の限定条件:等速運動または静止
光速不変:光の速度はどのような立場から見ても一定である、光よりも速い速度はあり得ない
同時性の破れ:異なる位置における同時性は、慣性系によって変わる(光速不変の原理による)
電車の中央から両壁に光を発射、電車内にいる観測者は両壁到達が同時
外のホームにいる観測者には準光速走行電車の両壁到達は同時ではない
時間の相対性:光の速度に近づくと、時間の流れはゆっくりになる
E=mc2(E:エネルギー、m:質量、c:光速)
質量をエネルギーに、エネルギーを質量に変えられる
1916年一般相対性理論
空間の相対性:重力の大きいところでは空間がゆがむ、大きな重力は光さえも曲げる
時間の相対性:重力の大きいところでは時間の流れはゆっくりになる
ファラデーの電磁誘導の法則 磁場が時間変化すると電場が生まれる
アンペールの法則 電流のまわりに磁場ができる
電場のガウスの法則 電場は+電荷から出て-電荷に吸い込まれる(+極/ー極)
磁場のガウスの法則 磁場は湧き出しも吸い込みも起こらない(常にNSペアの場)
アンペールの法則 電流のまわりに磁場ができる
電場のガウスの法則 電場は+電荷から出て-電荷に吸い込まれる(+極/ー極)
磁場のガウスの法則 磁場は湧き出しも吸い込みも起こらない(常にNSペアの場)
2枚の電極→交流電圧→変動する電場→変位電流→変動する磁場発生→変動する電場発生
→2枚の電極のすき間に電場と磁場が交互にあらわれる電磁波が発生し、
周辺に伝わっていく(光:電磁波:電子の往復運動で生じる)
→2枚の電極のすき間に電場と磁場が交互にあらわれる電磁波が発生し、
周辺に伝わっていく(光:電磁波:電子の往復運動で生じる)
電流(電子の流れ)は磁場を作る─┐
├→電磁場の振動=電磁波=光
磁場の変化は電流を作る─────┘
├→電磁場の振動=電磁波=光
磁場の変化は電流を作る─────┘
1900年プランクのエネルギー量子仮説 … (量子論の父)
溶鉱炉の鉄の温度と光の色の研究過程で光のエネルギーはとびとびであることを発見
黒体放射の研究からE=nhν(n=0,1,2,...)という数式を考案(h:プランク定数)
光のエネルギーの量子化、光=波長が連続的な波、を覆す?考え方
νという振動数(色)をもった光はhνの整数倍のエネルギーしか吸収も放射もできない
プランクは光とは言わず「振動子」と呼んでいた
光のエネルギーE,振動数ν,プランク定数h=6.62607015×10−34Js(Js=m2kg/s)
名言 … すべての物質は固有の振動数を持つ
1905年アインシュタインの光量子仮説
プランクのエネルギー量子仮説にヒントを得て光量子仮説を発表
光はエネルギーをもつ粒の集まり、光電効果→光は粒である
1905年は、アインシュタインが3つの大きな発見を発表した奇跡の年です
特殊相対性理論、光量子説に基づく光電効果の理論、ブラウン運動の理論、が
たった一人の物理学者によって提唱されました
100年目の2005年を世界物理年と制定、6月30日はアインシュタイン記念日
光がある一定の波長より短いと金属表面から電子が飛び出し、より長いと電子は飛び出さない
光のエネルギーE,振動数ν,プランク定数h,波長λ,仕事関数Wは金属内の電子取出エネルギー
光量子説:E=hν - W:放出電子のエネルギー最大値、光の粒子の振動数の量子化
・光量子の数を増やす →金属表面から飛び出す電子の数が増える
・光量子の振動数を増やす→金属表面から飛び出す電子のエネルギーが増える
1913年ボーアの原子構造の量子論 … (量子論の育ての親)
1911年、原子の中心には原子核があるとラザフォードが主張した … (原子核物理学の父)
入射したアルファ粒子の偏向からラザフォード散乱による原子核の発見
バルマーは水素ガスに電圧をかけると4色の光の波長に規則性があることに気づいた
ボーアの原子構造の量子論はバルマーの発見の話にヒントを得た
原子核の周りをまわる電子の軌道はとびとびの円軌道
電子の軌道半径Rは必ず次の条件を満たす(p:電子の運動量、n:任意の自然数)
2πRp=nh (n=1:基底状態、h:プランク定数) … ボーアの量子条件
電子が光子を放出・吸収して軌道を飛び移る際に生じる線スペクトルを説明できた
ボーアは量子条件の物理的な意味がわからなかったがその理由を考えたのがド・ブロイ
ラザフォードは後に陽子を発見、中性子の存在を予想、チャドウィックが中性子を発見
1924年ド・ブロイの物質波の提唱
ド・ブロイはボーアの電子のとびとび軌道の直観にボーアに代わってその理由を考えた
アインシュタインの光量子仮説を参考に物質の正体は波であるとして物質波と名付けた
電子の波が消えないためには電子の軌道の1周の長さは波の波長の整数倍に限られる
1周の長さが波長の整数倍、量子条件とは電子の波動性を示すものであった
ド・ブロイが提唱した物質波の概念を知ったシュレディンガーはこれは面白いと思った
1925年ハイゼンベルクの行列力学
量子力学の最初の厳密な定式化
行列力学は量子の世界を粒子の側面から離散的にとらえ、 … ハイゼンベルク
波動力学は量子の世界を波動の側面から連続的にとらえている … シュレディンガー
のちにディラックが発見した変換理論(一般的で抽象的な理論)により行列力学と
波動力学がつなげられ、一方の形式から他方の形式に変換できるようになりました
1925年シュレディンガー方程式の誕生
量子力学の基礎方程式:物質波の方程式
波の高さ(振幅)は複素数の縦軸、面積のような高さをもつ波
波の横軸は電子が存在する場所の広がりを表す
物質波の伝わり方を計算する方程式
波全体がある時刻における1個の電子の状態を表している
角振動数ω=2πν、Ψ=Acos(kx-ωt)+iAsin(kx-ωt)=Aei(kx-wt)
Ψ:座標表示の波動関数(プサイ)は確率振幅を表わす関数(波の高さ)
E:エネルギー固有値
H:ハミルトニアン、位置xと時間t依存運動量の関数=運動エネルギー+位置エネルギー
運動エネルギーT、位置エネルギーV、質量m、全エネルギーE=T+V=H
シュレディンガーの波動方程式は「観測前の波の状態」から、
観測によって「粒子が出現する座標(横軸)の確率(縦軸)」を予測する計算式のことです
1926年ボルンの主張した物質波の正体
シュレディンガーたちは物質波とはどんなものか示せず困っていた
ボルンは物質波の正体は確率の波であるという新たな仮説を主張した
電子のような小さな粒子の観測確率は波動関数の絶対値の2乗に比例する
物質波は確率の波、波動関数を電子位置の確率密度と考える解釈=統計的解釈
ある位置で電子が見出される確率はボルンの規則で与えられる(波動関数の確率解釈)
・波の高いところほど電子が発見される確率が高い
・見られる前の電子はさまざまな場所に広がっている(状態の重ね合わせ)
・見られた途端に波は収縮して電子は1箇所で発見される
P=|Ψ|2、確率P、波動関数Ψ、確率密度関数|Ψ|2 … ボルンの規則
1927年ハイゼンベルクの不確定性原理
ミクロの物質は位置と運動量を同時に確定することができない
電子の位置の測定誤差⊿xは光の波長λの程度
電子の運動量の測定誤差⊿pは⊿x⊿p≧h(プランク定数)/4π
⊿x⊿p≧ħ(ディラック定数)/2
位置と運動量の両者の不確かさを同時になくすのは不可能である
粒子の位置と運動量の測定に、必ず一定以上の不確かさが残ることを示す
運動量演算子pを波動関数ψに作用させ(px-xp)ψを計算すると次式([x,p]:q数)
px-xp=h/(2πi)、x:位置、p:運動量 … ディラックの量子条件
観測者効果ではなく、量子的なゆらぎによるとして改良された小澤の不等式が後日発表された
⊿x⊿p+σx⊿p+σp⊿x≧h/4π … ハイゼンベルクの式に2項追加
σx、σp:物体の位置と運動量が、測定前にもともと持っていた量子ゆらぎ(標準偏差)
1927年コペンハーゲン解釈
ボーアとハイゼンベルクは量子力学における正統的な解釈としても知られる
コペンハーゲン解釈をまとめた(名前の由来はボーア研究所の所在地)
標準解釈:ノイマンが1932年に行った定式化は
・量子系と観測者(観測装置)を分離する(2つの境界はどこに引いても良い)
・量子系の状態は、観測していないときはシュレディンガー方程式に従う
・観測により波動関数が収縮して、1つの測定値が得られる
・どの測定値が得られるかは確率的であり、ボルンの規則に従う
電子の運動はシュレディンガー方程式を満たす波動関数Ψ(r,t)で記述される
波動関数は一般に複素関数で、空間的に広がりを持ち、干渉や回折などの波に特有な性質を現す
電子の位置を実験的に観測した場合には電子はある一点に見出され、広がりを持たない
位置rの周りの微小体積drに電子が発見される確率Pは |Ψ(r,t)|2drに比例する
電子は空間のどこかに存在するはず → ∫Pdr=1 規格化された波動関数
規格化:全空間での積分が1になるような比例定数を見つけるだけの作業、波動関数の調整
電子は波であり粒子である
・波と粒子の両方の性質を取り入れて新しい量子力学の理論が作れる
・電子は観測されないときだけ、波として広がっている
・さまざまな場所にいる状態が重ね合わさってる、状態の重ね合わせ
・電子を観測すると電子の波は一瞬で収縮する
・状態の重ね合わせが解除されて、波が一点に集まり電子は粒子の姿に見える
・波の収縮場所は波動関数の確率解釈で決まる
1928年ディラック方程式
シュレディンガー方程式を特殊相対論の要請を満足するように修正した方程式
ディラックは、この方程式によりコペンハーゲン解釈の問題点を解消させた
溶鉱炉の鉄の温度と光の色の研究過程で光のエネルギーはとびとびであることを発見
黒体放射の研究からE=nhν(n=0,1,2,...)という数式を考案(h:プランク定数)
光のエネルギーの量子化、光=波長が連続的な波、を覆す?考え方
νという振動数(色)をもった光はhνの整数倍のエネルギーしか吸収も放射もできない
プランクは光とは言わず「振動子」と呼んでいた
光のエネルギーE,振動数ν,プランク定数h=6.62607015×10−34Js(Js=m2kg/s)
名言 … すべての物質は固有の振動数を持つ
1905年アインシュタインの光量子仮説
プランクのエネルギー量子仮説にヒントを得て光量子仮説を発表
光はエネルギーをもつ粒の集まり、光電効果→光は粒である
1905年は、アインシュタインが3つの大きな発見を発表した奇跡の年です
特殊相対性理論、光量子説に基づく光電効果の理論、ブラウン運動の理論、が
たった一人の物理学者によって提唱されました
100年目の2005年を世界物理年と制定、6月30日はアインシュタイン記念日
光がある一定の波長より短いと金属表面から電子が飛び出し、より長いと電子は飛び出さない
光のエネルギーE,振動数ν,プランク定数h,波長λ,仕事関数Wは金属内の電子取出エネルギー
光量子説:E=hν - W:放出電子のエネルギー最大値、光の粒子の振動数の量子化
c
c=λν から E=h─ - W 左式によりある一定の波長より短い必要性がわかる
λ
光は量子化された粒子であるとすると光電効果を説明できるc=λν から E=h─ - W 左式によりある一定の波長より短い必要性がわかる
λ
・光量子の数を増やす →金属表面から飛び出す電子の数が増える
・光量子の振動数を増やす→金属表面から飛び出す電子のエネルギーが増える
1913年ボーアの原子構造の量子論 … (量子論の育ての親)
1911年、原子の中心には原子核があるとラザフォードが主張した … (原子核物理学の父)
入射したアルファ粒子の偏向からラザフォード散乱による原子核の発見
バルマーは水素ガスに電圧をかけると4色の光の波長に規則性があることに気づいた
ボーアの原子構造の量子論はバルマーの発見の話にヒントを得た
原子核の周りをまわる電子の軌道はとびとびの円軌道
電子の軌道半径Rは必ず次の条件を満たす(p:電子の運動量、n:任意の自然数)
2πRp=nh (n=1:基底状態、h:プランク定数) … ボーアの量子条件
電子が光子を放出・吸収して軌道を飛び移る際に生じる線スペクトルを説明できた
ボーアは量子条件の物理的な意味がわからなかったがその理由を考えたのがド・ブロイ
ラザフォードは後に陽子を発見、中性子の存在を予想、チャドウィックが中性子を発見
1924年ド・ブロイの物質波の提唱
ド・ブロイはボーアの電子のとびとび軌道の直観にボーアに代わってその理由を考えた
アインシュタインの光量子仮説を参考に物質の正体は波であるとして物質波と名付けた
電子の波が消えないためには電子の軌道の1周の長さは波の波長の整数倍に限られる
1周の長さが波長の整数倍、量子条件とは電子の波動性を示すものであった
ド・ブロイが提唱した物質波の概念を知ったシュレディンガーはこれは面白いと思った
h
2πR=nλ よって pλ=h から λ=── (m:質量、v:速度、p=mv)
mv
2πR=nλ よって pλ=h から λ=── (m:質量、v:速度、p=mv)
mv
1925年ハイゼンベルクの行列力学
量子力学の最初の厳密な定式化
行列力学は量子の世界を粒子の側面から離散的にとらえ、 … ハイゼンベルク
波動力学は量子の世界を波動の側面から連続的にとらえている … シュレディンガー
のちにディラックが発見した変換理論(一般的で抽象的な理論)により行列力学と
波動力学がつなげられ、一方の形式から他方の形式に変換できるようになりました
1925年シュレディンガー方程式の誕生
量子力学の基礎方程式:物質波の方程式
波の高さ(振幅)は複素数の縦軸、面積のような高さをもつ波
波の横軸は電子が存在する場所の広がりを表す
物質波の伝わり方を計算する方程式
波全体がある時刻における1個の電子の状態を表している
角振動数ω=2πν、Ψ=Acos(kx-ωt)+iAsin(kx-ωt)=Aei(kx-wt)
∂Ψ
iħ──=HΨ=EΨ … 時間依存型線型偏微分方程式
∂t
i:虚数単位、h:プランク定数、ħ=(h/2π):ディラック定数iħ──=HΨ=EΨ … 時間依存型線型偏微分方程式
∂t
Ψ:座標表示の波動関数(プサイ)は確率振幅を表わす関数(波の高さ)
E:エネルギー固有値
H:ハミルトニアン、位置xと時間t依存運動量の関数=運動エネルギー+位置エネルギー
運動エネルギーT、位置エネルギーV、質量m、全エネルギーE=T+V=H
ħ2 ∂2
H=-──・──+V (x,t)
2m ∂x2
シュレディンガーは彼のアプローチが行列力学と等価であることを示すことができたH=-──・──+V (x,t)
2m ∂x2
シュレディンガーの波動方程式は「観測前の波の状態」から、
観測によって「粒子が出現する座標(横軸)の確率(縦軸)」を予測する計算式のことです
1926年ボルンの主張した物質波の正体
シュレディンガーたちは物質波とはどんなものか示せず困っていた
ボルンは物質波の正体は確率の波であるという新たな仮説を主張した
電子のような小さな粒子の観測確率は波動関数の絶対値の2乗に比例する
物質波は確率の波、波動関数を電子位置の確率密度と考える解釈=統計的解釈
ある位置で電子が見出される確率はボルンの規則で与えられる(波動関数の確率解釈)
・波の高いところほど電子が発見される確率が高い
・見られる前の電子はさまざまな場所に広がっている(状態の重ね合わせ)
・見られた途端に波は収縮して電子は1箇所で発見される
P=|Ψ|2、確率P、波動関数Ψ、確率密度関数|Ψ|2 … ボルンの規則
1927年ハイゼンベルクの不確定性原理
ミクロの物質は位置と運動量を同時に確定することができない
電子の位置の測定誤差⊿xは光の波長λの程度
電子の運動量の測定誤差⊿pは⊿x⊿p≧h(プランク定数)/4π
⊿x⊿p≧ħ(ディラック定数)/2
位置と運動量の両者の不確かさを同時になくすのは不可能である
粒子の位置と運動量の測定に、必ず一定以上の不確かさが残ることを示す
運動量演算子pを波動関数ψに作用させ(px-xp)ψを計算すると次式([x,p]:q数)
px-xp=h/(2πi)、x:位置、p:運動量 … ディラックの量子条件
観測者効果ではなく、量子的なゆらぎによるとして改良された小澤の不等式が後日発表された
⊿x⊿p+σx⊿p+σp⊿x≧h/4π … ハイゼンベルクの式に2項追加
σx、σp:物体の位置と運動量が、測定前にもともと持っていた量子ゆらぎ(標準偏差)
1927年コペンハーゲン解釈
ボーアとハイゼンベルクは量子力学における正統的な解釈としても知られる
コペンハーゲン解釈をまとめた(名前の由来はボーア研究所の所在地)
標準解釈:ノイマンが1932年に行った定式化は
・量子系と観測者(観測装置)を分離する(2つの境界はどこに引いても良い)
・量子系の状態は、観測していないときはシュレディンガー方程式に従う
・観測により波動関数が収縮して、1つの測定値が得られる
・どの測定値が得られるかは確率的であり、ボルンの規則に従う
電子の運動はシュレディンガー方程式を満たす波動関数Ψ(r,t)で記述される
波動関数は一般に複素関数で、空間的に広がりを持ち、干渉や回折などの波に特有な性質を現す
電子の位置を実験的に観測した場合には電子はある一点に見出され、広がりを持たない
位置rの周りの微小体積drに電子が発見される確率Pは |Ψ(r,t)|2drに比例する
電子は空間のどこかに存在するはず → ∫Pdr=1 規格化された波動関数
規格化:全空間での積分が1になるような比例定数を見つけるだけの作業、波動関数の調整
電子は波であり粒子である
・波と粒子の両方の性質を取り入れて新しい量子力学の理論が作れる
・電子は観測されないときだけ、波として広がっている
・さまざまな場所にいる状態が重ね合わさってる、状態の重ね合わせ
・電子を観測すると電子の波は一瞬で収縮する
・状態の重ね合わせが解除されて、波が一点に集まり電子は粒子の姿に見える
・波の収縮場所は波動関数の確率解釈で決まる
1928年ディラック方程式
シュレディンガー方程式を特殊相対論の要請を満足するように修正した方程式
ディラックは、この方程式によりコペンハーゲン解釈の問題点を解消させた
| 量子力学的な現象 | コペンハーゲン解釈 |
|---|---|
| 波と粒子の二重性 | 量子は波の性質と粒子の性質をもつ、同時両有は排除するが相補性がある |
| 不連続性 | 粒子が吸収・放出するエネルギーはとびとび、エネルギーは光子のような量子 |
| 不確定性原理 | 観測が粒子の状態を乱すため、位置と運動のどちらかの正確性が失われる |
| 確率解釈 | 波動関数の絶対値の2乗は粒子の存在確率を示す |
| 重ね合わせ | 観測する迄は波動関数がとり得るさまざまな状態にある |
| トンネル効果 | 反射と透過の重ね合わせ、量子がエネルギーの壁を確率的にすり抜ける現象 |
| 排他原理 | 物質を構成する粒子は、同じ種類が1つの場所に同時には存在しえない |
| 非局所性 | 量子もつれ状態が局所性を破るEPRパラドックス、量子テレポーテーション |
ミクロの世界のルールが判明:ボーア、ボルン、ハイゼンベルグが主張
ルールその1:ミクロの物質のふるまいは、ミクロの物質の正体を波と考えれば説明できる
ルールその2:ミクロの物質のふるまいは、確率的にしかわからない
プランク、アインシュタイン、ド・ブロイ、シュレディンガーは、ルールその2に絶対反対
ルールその1:ミクロの物質のふるまいは、ミクロの物質の正体を波と考えれば説明できる
ルールその2:ミクロの物質のふるまいは、確率的にしかわからない
プランク、アインシュタイン、ド・ブロイ、シュレディンガーは、ルールその2に絶対反対
|
アインシュタインら3名の名前の頭文字からEPRと称し、量子もつれ(絡み合い)状態の非局所 性を指し、量子の状態は特定の状態にはなく、誰かが観測した瞬間にその状態が決まると考える すると、関係しあう2個の量子は、一方の量子の状態が決定すると、はじめて他方の状態も決ま るということが起こり、これらの量子をはるか遠方に引き離してもそれは同じだという |
|
量子エンタングルメント(量子もつれ)は、観測された一方の粒子から、 もう一方に情報が(光速を超えて)伝達されることではなく、 互いに離れた粒子の性質が、セットで決まっていることだと考えられている |
|
送りたい光子状態:光子Xの状態 … 例:水平偏光と垂直偏光の重ね合わせ
↓転送光子準備:EPR対(量子もつれの関係)の光子Aと光子Bの生成 ↓ 光子Bの転送 ↓ 光子Xと光子Aは量子もつれ状態でベル測定(光子Xと光子Aを同時に観測) ↓ 測定結果の古典通信:2ビット値(00,01,10,11)のいずれかを送信(光子Xのズレの補正) ↓ 受信した測定結果から光子Bの偏光操作(補正後の光子B=光子X) ↓ 光子Xの復元(状態情報の復元)
ベル測定:ハーフミラーを用いたビームスプリッターでは、2光子の反射と透過の2種の
組み合わせで4通りあり、この状態を測定することをベル測定と言う 2光子は同時にビームスプリッターに達するようにしなくてはいけない |
量子もつれを生かし、物質の状態をそっくり迅速に遠くへ移すこと
もつれた粒子対(EPR対)の片方を送り手が、他方を受け手がもつ
送り手の粒子を第三の粒子ともつれさせ、この観測結果を通常通信で
受け手に知らせると、その情報を使って受け手の粒子に第三の粒子の
状態を再現できる
もつれた粒子対(EPR対)の片方を送り手が、他方を受け手がもつ
送り手の粒子を第三の粒子ともつれさせ、この観測結果を通常通信で
受け手に知らせると、その情報を使って受け手の粒子に第三の粒子の
状態を再現できる
量子の状態を完全に表現するには波の性質と粒子の性質のどちらも必要
ボーアはこの性質を「相補性」と呼ぶ
位置を特定すると速度が曖昧になり速度を特定すると位置が曖昧になる
という不確定性原理も、相補性の現れであるとされる
ボーアはこの性質を「相補性」と呼ぶ
位置を特定すると速度が曖昧になり速度を特定すると位置が曖昧になる
という不確定性原理も、相補性の現れであるとされる
波の収縮は起こらず、観測された瞬間の波の収縮をシュレディンガー方程式から数学的に導く
ことは原理的に不可能であるとジョン・フォン・ノイマンが証明した
状態の重ね合わせでなく多数の世界の同時存在として説明し直したが多数の世界の同時存在の
証明はできていない(分岐した世界は孤立しているとされている)
ことは原理的に不可能であるとジョン・フォン・ノイマンが証明した
状態の重ね合わせでなく多数の世界の同時存在として説明し直したが多数の世界の同時存在の
証明はできていない(分岐した世界は孤立しているとされている)
ディラックが創始した理論、場の量子化、場の量子論の原型はハイゼンベルクとパウリが創った
従来の量子力学では素粒子が生成・消滅しないものと考え、その座標や運動量を演算子で表すが、
空間の各点での場や粒子数を演算子とし(場の量子化)、その変化を量子論的に追求する理論
これにより素粒子の性質(生成・消滅も含めて)、それらの間の相互作用が統一的に記述される
波または粒子が実在するのではなく、場が振動することで波や粒子に見える
振動が広い範囲で連動すると波に見え、長い時間保持する振動は粒子に見える
力の相互作用はゲージ粒子のキャッチボールではなく、波でもあり粒子でもあるような場の振動
量子電磁力学、量子色力学、電弱相互作用の理論
従来の量子力学では素粒子が生成・消滅しないものと考え、その座標や運動量を演算子で表すが、
空間の各点での場や粒子数を演算子とし(場の量子化)、その変化を量子論的に追求する理論
これにより素粒子の性質(生成・消滅も含めて)、それらの間の相互作用が統一的に記述される
波または粒子が実在するのではなく、場が振動することで波や粒子に見える
振動が広い範囲で連動すると波に見え、長い時間保持する振動は粒子に見える
力の相互作用はゲージ粒子のキャッチボールではなく、波でもあり粒子でもあるような場の振動
量子電磁力学、量子色力学、電弱相互作用の理論
量子論に基づいて宇宙の始まりを考える
ビッグバン宇宙(確実に正しい)
ジョルジュ・ルメートルが宇宙膨張説を発表
エドウィン・ハッブルが膨張宇宙を発見、遠くの銀河ほど遠ざかる、空間自体が膨張
ガモフの火の玉宇宙論(ビッグバン理論)
宇宙マイクロ背景輻射(CMB)の発見:宇宙が昔高温・高密度であった痕跡
WMAP・Planck衛星が描く宇宙創成30万年後の宇宙地図(宇宙の晴れ上がり面)
宇宙マイクロ波背景放射の温度ゆらぎはビッグバン理論の予言と一致
ビッグバン以前(理論的予想)
真空は時空の泡状態で粒子・反粒子が生成・消滅している状態
真空のエネルギーを想定したインフレーション理論など様々なインフレーション理論
ビレンケンの無からの宇宙創成論:量子トンネル効果によって無から極小な宇宙が生まれた
宇宙が虚数の時間に生まれたというホーキングの無境界仮説
誕生直後の宇宙の半径はプランクサイズ→インフレーション→ビッグバン宇宙の誕生
ビッグバン宇宙(確実に正しい)
ジョルジュ・ルメートルが宇宙膨張説を発表
エドウィン・ハッブルが膨張宇宙を発見、遠くの銀河ほど遠ざかる、空間自体が膨張
ガモフの火の玉宇宙論(ビッグバン理論)
宇宙マイクロ背景輻射(CMB)の発見:宇宙が昔高温・高密度であった痕跡
WMAP・Planck衛星が描く宇宙創成30万年後の宇宙地図(宇宙の晴れ上がり面)
宇宙マイクロ波背景放射の温度ゆらぎはビッグバン理論の予言と一致
ビッグバン以前(理論的予想)
真空は時空の泡状態で粒子・反粒子が生成・消滅している状態
真空のエネルギーを想定したインフレーション理論など様々なインフレーション理論
ビレンケンの無からの宇宙創成論:量子トンネル効果によって無から極小な宇宙が生まれた
宇宙が虚数の時間に生まれたというホーキングの無境界仮説
誕生直後の宇宙の半径はプランクサイズ→インフレーション→ビッグバン宇宙の誕生
人間スケール … 古典物理学:力学、電磁気学、熱力学、物性物理・化学、天体物理学・・・
ミクロの世界 … 量子物理学:核物理、素粒子物理、確率解釈、場の量子論、相対論的量子論
宇宙スケール … 相対性理論:4次元時空、重力理論、光速不変、E=mc2
上記3つの物理学領域は一部重なり合っている
ミクロの世界 … 量子物理学:核物理、素粒子物理、確率解釈、場の量子論、相対論的量子論
宇宙スケール … 相対性理論:4次元時空、重力理論、光速不変、E=mc2
上記3つの物理学領域は一部重なり合っている
「量子」は物質ではなく、波動性を持った状態で「エネルギー」として存在していて、
外部からの観測のエネルギーを受けた瞬間に一点に収縮した「粒子」となって出現します
この「エネルギー状態の量子」が外部からの観測のエネルギーを受けた瞬間に一点に収縮
した粒子となって出現する現象のことを波動関数の収縮(=デコヒーレンス)と呼びます
「量子レベルの物質」は実際に観測されたものだけが物質化しており、観測されない限り
「そこにあるかもしれないし、ないのかもしれない」
という『可能性の波』として存在しているということです
「見ていないものは存在すらしないのかもしれない」というような話です
コペンハーゲン解釈は観測できない観測以前の状態を推測して自然界の法則を導こうとする
論理的な仮説でしかなく、そもそもどれが絶対的に正しいという話でもありません
人間は、量子の振る舞いに関するその考え方を正しいか、
間違っているかと判断する手段を持ち合わせていない。
そのような考え方をしたところで実際には何の矛盾も
起きないからこそ採用する。
量子力学は人間の脳が理解できる限界を超えた現象をあるがまま法則として受け入れていると
していますが、とりあえずはそうしておいて、いつの日にか壁を越えて終着駅にたどり着ける
こともかすかに期待しつつ、量子論のおかげで成り立つ最新技術の製品を手にしながら自らを
納得させておきます
外部からの観測のエネルギーを受けた瞬間に一点に収縮した「粒子」となって出現します
この「エネルギー状態の量子」が外部からの観測のエネルギーを受けた瞬間に一点に収縮
した粒子となって出現する現象のことを波動関数の収縮(=デコヒーレンス)と呼びます
「量子レベルの物質」は実際に観測されたものだけが物質化しており、観測されない限り
「そこにあるかもしれないし、ないのかもしれない」
という『可能性の波』として存在しているということです
「見ていないものは存在すらしないのかもしれない」というような話です
コペンハーゲン解釈は観測できない観測以前の状態を推測して自然界の法則を導こうとする
論理的な仮説でしかなく、そもそもどれが絶対的に正しいという話でもありません
人間は、量子の振る舞いに関するその考え方を正しいか、
間違っているかと判断する手段を持ち合わせていない。
そのような考え方をしたところで実際には何の矛盾も
起きないからこそ採用する。
量子力学は人間の脳が理解できる限界を超えた現象をあるがまま法則として受け入れていると
していますが、とりあえずはそうしておいて、いつの日にか壁を越えて終着駅にたどり着ける
こともかすかに期待しつつ、量子論のおかげで成り立つ最新技術の製品を手にしながら自らを
納得させておきます
半導体と量子論 … 半導体の仕組みはバンド理論によって説明できる
光のエネルギーを電気に変換する、電気のエネルギーを光に変換する、
電気の流れを制御できる
情報機器、家電製品、LED電球
超流動と超伝導 … 液体ヘリウムの超流動はボース=アインシュタイン凝縮で説明される
超流動が電子で起こると超伝導と呼ばれ、クーパー対を作った電子は
ボース粒子としてふるまう
超伝導体だけで作った回路は電気抵抗がなくなり永久電磁石になる
リニアモーター
量子コンピュータ … 量子論的な状態の重ね合わせの原理にもとづく量子ビットを用いる
量子もつれ … 量子テレポーテーション
光のエネルギーを電気に変換する、電気のエネルギーを光に変換する、
電気の流れを制御できる
情報機器、家電製品、LED電球
超流動と超伝導 … 液体ヘリウムの超流動はボース=アインシュタイン凝縮で説明される
超流動が電子で起こると超伝導と呼ばれ、クーパー対を作った電子は
ボース粒子としてふるまう
超伝導体だけで作った回路は電気抵抗がなくなり永久電磁石になる
リニアモーター
量子コンピュータ … 量子論的な状態の重ね合わせの原理にもとづく量子ビットを用いる
量子もつれ … 量子テレポーテーション
TOP|未来光円錐|TOP
3次元空間+1次元時間=4次元時空を表現する「光円錐」(の上半分)
3次元空間+1次元時間=4次元時空を表現する「光円錐」(の上半分)
時間
・←──────────────・──────────────→・
\ ↑ /
\ │ /
\ 原点から出発 │ 原点から出発 /
\ した光が到達 │ した光が到達 /
光\ し得る範囲 │ し得る範囲 /光
速\ │ /速
の\ │ /の
傾\ │時 /傾
き\ │間 /き
\ │の /
\ │流 /
\ │れ /
\ │ /
\ │ /
未来\│/未来
空間・←──────────────●──────────────→・空間
事象の地平線(光速の壁)の外側は認識できない領域 = 存在しない?
・←──────────────・──────────────→・
\ ↑ /
\ │ /
\ 原点から出発 │ 原点から出発 /
\ した光が到達 │ した光が到達 /
光\ し得る範囲 │ し得る範囲 /光
速\ │ /速
の\ │ /の
傾\ │時 /傾
き\ │間 /き
\ │の /
\ │流 /
\ │れ /
\ │ /
\ │ /
未来\│/未来
空間・←──────────────●──────────────→・空間
事象の地平線(光速の壁)の外側は認識できない領域 = 存在しない?
<宇宙の根本原理>
|TOP|
エントロピー増大の法則:自発変化の方向を示した法則であり、
その方向とは不可逆的な無秩序化だということです エントロピーが低い状態:整然、エネルギーの質=高い(利用しやすい) エントロピーが高い状態:乱雑、エネルギーの質=低い(利用しにくい)
エントロピー増大リスク:超新星爆発、小天体の衝突、気候温暖化
宇宙の歴史の根源的な流れ → エントロピー減少の方向に進んでいる
エネルギーの凝集により物質(光子→素粒子→核子→原子核→原子)が生まれる 原子の凝集→核融合→太陽→ダストの凝集→惑星→有機物→生命の誕生 太陽→植物の光合成→酸素→多細胞生物→生物の陸上進出→人類の出現
時間スケール(秒)の各行の時間を空間スケール(m)の各行の長さにするには光速(3×108m/秒)をかければ良い
時間スケールで示された時間内に光の信号が到達できる距離が空間スケールで示される長さになっています 時間スケールの時間の記載がない行は空間スケールの長さ(m)に光速の逆数(3.33×10-9秒/m)をかければ良い 人間スケールで見てみると、大きさに関しては大きからず小さからず図のなかほどで中間的な存在なのです 時間に関しては人間の寿命が宇宙の年齢と同じくらい長いのには驚かされます
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}